|
ACADEMIA ROMÂNĂ |
||||||||||||||||||||||||
|
Cursul Postuniversitar de studii avansate “Realizari si perspective in Biologie”, ediţia 2008 PROGRAM 8:30 – 9:00 – Înregistrarea participanţilor 9:00 – 9:15 – Deschiderea oficială 9:15 – 10:00 - Dr. Anca Păunescu - Institutul de Biologie Bucureşti Răspunsul biodiversităţii la schimbările climatice la nivelul principalelor categorii de ecosisteme 10:00 – 10:45 - Dr. Nicolae Mirancea - Institutul de Biologie Bucureşti Rolul interactiunilor gene-mediu in etiologia cancerului uman 10:45 – 11:00 Pauză de cafea 11:00 – 11:45 – Prof. Dr. Ioan Ardelean - Institutul de Biologie Bucureşti Contribuţia unor procariote marine la consumarea dioxidului de carbon si metanului 11:45 – 12:30- Dr. Aurelia Brezeanu - Institutul de Biologie Bucureşti Stresul oxidativ indus de factorii de mediu, element perturbator major al proceselor de dezvoltare la plante 12:30 – 13:30 Pauză de masă 13:30 – 14:15 - Dr. Cristina Sandu - Institutul de Biologie Bucureşti Impactul antropic asupra ecosistemelor acvatice – efecte şi feed-back 14:15 – 15:00 – Dr. Dorina Purice – Institutul de Biologie Bucureşti Bioindicatorii şi semnificaţia scalei spaţio-temporale în aspectele legate de schimbările climatice
9:00 – 9:45 – Dr. Liliana Oromulu - Institutul de Biologie Bucureşti 9:45 – 10:30 - Dr. Sorin Stefănuţ - Institutul de Biologie Bucureşti Impactul potenţial al schimbărilor climatice asupra ecosistemelor alpine 10:30 – 11:15 - Dr. Sanda Maican - Institutul de Biologie Bucureşti Impactul schimbărilor climatice asupra insectelor 11:15 – 11:30 Pauză de cafea
11:30 – 12:15 - Dr. Viorica Honciuc - Institutul de Biologie Bucureşti Influenţa factorilor de mediu asupra biodiversităţii faunei de sol din ecosistemele forestiere 12:15 – 13:00- Dr. Laura Parpală, Dr. Victor Zinevici, Dr. Doina Ionică, Mirela Moldoveanu, Larisa Florescu – Institutul de Biologie Bucureşti Biodiversitate, stabilitate şi productivitate în condiţii ecologice naturale şi de impact antropic 13:00 – 14:00 Pauză de masă 14:00 – 14:45 - Dr. Marilena Onete - Institutul de Biologie Bucureşti Impactul factorilor de mediu asupra biodiversităţii urbane 14:45 – 15:30 - Dr. Carmen Mădălina Cişmaşiu - Institutul de Biologie Bucureşti
9:00 - 9:45 – Dr. Petruta Oancea - Fac. Chimie, Univ. din Bucuresti Poluarea apelor şi metodele adecvate pentru distrugerea poluanţilor 9:45-10:30 - Prof. dr. Ioana Gomoiu - Institutul de Biologie Bucureşti Impactul modificărilor climatice asupra fungilor 10:30 -11:10 - Dr. Mădălin Enache - Institutul de Biologie Bucureşti Halobacteriile şi implicaţiile lor în formarea evaporitelor 11:10 -11:30 - Înmânarea diplomelor; închiderea lucrărilor
REZUMATE Răspunsul biodiversităţii la schimbările climatice la nivelul principalelor categorii de ecosisteme Anca Păunescu Institutul de Biologie Bucureşti, e-mail: ancuta_paun@yahoo.com Sistemul climatic cuprinde ca subsisteme principale, atmosfera, oceanul, suprafaţa uscatului, biosfera şi criosfera. Natura diferită a acestor subsisteme care sunt în permanentă interrelaţie, generează variabilitatea climatică. Schimbările climatice sunt rezultatul acţiunii simultane a factorilor interni (modificări care apar în interiorul sistemului climatic sau datorită interacţiunilor dintre subsistemele sale), externi naturali (variaţii ale energiei solare, erupţii vulcanice, variaţii ale parametrilor orbitali ai Pamântului) şi externi antropogeni, rezultaţi din activităţile umane (schimbarea compoziţiei atmosferei ca urmare a creşterii concentraţiei gazelor care produc efectul de seră). Unele gaze atmosferice au efect protector asupra planetei prin absorbţia, dar mai ales prin emisia de radiaţii infraroşii. În lipsa acestor gaze suprafaţa planetei s-ar răci şi ar deveni nelocuibilă. Nivelul natural al acestor gaze (bioxid de carbon, metan, ozon, bioxid de azot, etc.) a fost suplimentat dramatic în ultimii o sută de ani, de emisii rezultate în urma activităţilor umane, ca de exemplu, arderea combustibililor fosili, diverse activiăţi industriale şi practicarea agriculturii intensive. În consecinţă, au apărut dezechilibre între absorbţia şi emisia de radiaţii infraroşii care au condus la o încălzire a suprafeţei terestre, aşa-numitul efect de seră. Din 1896, când a fost măsurat pentru prima dată de către S.Arrhenius, şi până în prezent, efectul de seră a determinat o creştere medie globală a temperaturii atmosferice cu 0.6°C. Conform ultimelor estimări, creşterea temperaturii atmosferice a fost mai accentuată în America de Nord, în zona ecuatorială şi în Asia (0.8°C). Chiar dacă poate părea redusă, această creştere a temperaturilor medii, a condus la schimbări climatice semnificative, fiind afectate la scară globală, toate ecosistemele. Ecosistemele polare s-au modificat vizibil în ultimii 100 de ani. De exemplu, pădurea boreală înaintează în Alaska, spre nord, cu 100 km pentru fiecare grad Celsius creştere de temperatură, concomitent cu reducerea cu 40% a calotei glaciare arctice, doar în ultimii 30 de ani; populaţiile de urşi polari şi de pinguini imperiali s-au redus la jumătate. În ecosistemele agricole, schimbările climatice precum şi capacitatea redusă de adaptare la stresul hidric şi biotic a plantelor cultivate, a redus productivitatea agricolă la aproximativ o treime, având consecinţe dramatice, mai ales la nivelul ţărilor mai puţin dezvoltate, cu economie preponderent agrară. În acelaşi timp, agricultura influenţează direct schimbărille climatice. Se apreciază că aproximativ 20% din emisiile de gaze de seră sunt rezultate din activităţi agricole (utilizarea de amendamente azotoase, creşterea animalelor rumegătoare, incendierea resturilor vegetale, etc.). La nivelul ecosistemelor aride (deşert, savană, stepă, etc.) se consideră că speciile caracteristice au atins deja pragul critic de adaptabilitate şi sunt într-un declin considerabil. Se poate vorbi despre teritorii extinse cu grad înalt de vulnerabilitate, aşa-numitele hot spot, cum este de exemplu, aria denumită Succulent Karoo, cu cea mai mare diversitate de Crassulaceae şi Cactaceae din lume, 69% dintre specii fiind endemice. În ecosistemele forestiere atât diversitatea floristică cât şi cea faunistică sunt ameninţate; în prezent, se apreciază că peste 50% dintre primate şi 9% dintre speciile de plante lemnoase sunt în pericol de dispariţie. Schimbările climatice au impact considerabil şi asupra ecosistemelor alpine, provocând retragerea şi uneori chiar dispariţia de specii. De exemplu, în Munţii Alpi multe dintre speciile de plante alpine au migrat ascendent înaintând aproximativ 4 metri la fiecare 10 ani iar unele specii care populau numai vârfurile muntoase, deja au dispărut. Şi ecosistememele acvatice sunt afectate de încălzirea globală, deopotrivă cele dulcicole şi şi cele marine. Se apreciază că în ultimii 20 de ani, aproximativ 20% dintre speciile de peşti dulcicoli au devenit ameninţaţi cu dispariţia, de asemenea, majoritatea mamiferelor marine sunt pe listele speciilor periclitate. Marea barieră de corali, considerată padurea tropicală a mărilor deoarece adăposteşte peste 25% dintre speciile marine, este serios ameninţată cu dispariţia datorită accentuării fenomenelor de degradare a coralilor (albire), mult mai frecvente în ultimii 50 de ani, consecinţă directă a încălzirii climatice. Acest declin general al ecosistemelor poate fi redus prin adoptarea de măsuri prin care să se diminueze emisiile de gaze de seră. Între acestea, cel mai des discutate sunt: reducerea consumurilor energetice, utilizarea energiilor şi a combustibililor neconvenţionali, aplicarea de tehnici agricole nepoluante, etc. Aceste măsuri trebuie luate cât mai rapid deoarece chiar dacă în final se va obţine diminuarea şi stabilizarea concentraţiei gazelor care produc efectul de seră, datorită inerţiei sistemului climatic, pentru o perioadă de timp determinată, clima va continua să se încălzească.
ROLUL INTERACŢIUNILOR GENE-MEDIU ÎN ETIOLOGIA CANCERULUI N. Mirancea, Dorina Mirancea Institutul de Biologie Bucureşti CANCERUL – este un termen generic care defineşte un grup de boli care sunt iniţiate şi se manifestă clinic atunci când unele celule ale corpului încep să se multiplice şi să migreze necontrolat. Corpul nostru este o comunitate de celule, în care fiecare celulă ocupă un loc potrivit pentru funcţia sa. Cu excepţia celulelor sanguine care sunt abilitate să patruleze prin întregul organism pentru asanarea microbilor invadatori şi a ţesuturilor alterate, celulele normale stau în ţesutul din care acestea fac parte. Totuşi, celulele canceroase violează această regulă şi trec agresiv în ţesuturile vecine. Celula ca entitate genetică şi sistem biologic deschis, în limita normei individuale de reactivitate, răspunde adecvat stimulilor extracelulari care, via receptori celulari şi molecule transductoare activează şi represează succesiv diferite segmente ale programului genetic cu “scopul” de a asigura supravieţuirea şi multiplicarea celulei. Citodiferenţierea şi realizarea unor structuri supracelulare care să funcţioneze armonios în cadrul unui organism, necesită realizarea unor cicluri celulare ordonate spaţio-temporal. Există mecanisme care reglează foarte precis atât ritmul proliferării cât şi locaţia temporal-vectorială a diferitelor tipuri de celule. Organismele eukariote superioare au o durată limitată de viaţă ca urmare a faptului că celulele care le compun realizează un număr limitat de cicluri celulare controlate de mecanisme intracelulare care răspund influenţelor exercitate de celulele vecine şi/sau de mediul extracelular. O excepţie o constituie celulele canceroase care se divid necontrolat şi care manifestă caracteristici de celule dediferenţiate capabile să se înmulţească nelimitat, ceea ce este letal pentru individul în care acestea apar. Transformarea malignă este un proces multistadial care, în principal se poate realiza prin acumularea unor mutaţii la nivelul a două clase de gene care au rol major în declanşarea cancerului: (1) proto-oncogene şi (2) gene supresoare de tumori. În configuraţia lor normală (1) şi (2) controlează ciclul celular de creştere, diviziune celulară, citodiferenţiere şi dezvoltare embrionară. Majoritatea proto-oncogenelor codifică proteine care transmit semnale venite din afara celulei, care via receptori plasmalemali generează semnale stimulatorii. Genele supresoare de tumori au efecte contrare, inhibitorii. Oncogenele sunt versiuni modificate (alterate) ale genelor obişnuite (proto-oncogene) benigne, care în celulele normale codifică proteine utilizate de celulă în realizarea unor structuri şi funcţii normale. Oncogenele au fost identificate prima dată la virusuri, dar există opinia că acestea provin din gene celulare care au fost preluate şi încorporate în genomul viral în timpul infectării celulelor. Celula malignă provine dintr-o celulă iniţial normală, care prin acumularea unor mutaţii la nivelul proto-oncogenelor şi/sau al genelor supresoare de tumori se transformă într-un fenotip nefast care va prolifera anarhic (necontrolat) şi va introduce dezordine în subsistemele organismului. Constituirea statutului de malignitate a unei celule şi formarea unei mase tumorale primare presupune ca celula respectivă să se dividă pentru ca alterările genetice achiziţionate să se multiplice în numeroşi descendenţi clonali care vor manifesta comportamente speciale. Cancerul se instalează atunci când una sau mai multe gene care controlează rata proliferării unui anumit tip celular şi a poziţionării în organism a celulelor rezultate prin multiplicare sunt alterate, ceea ce permite creşterea aberantă a celulelor respective şi, cel mai adesea invadarea ţesuturilor înconjurătoare sau distant localizate. Sunt peste 200 de tipuri de cancer, care într-o clasificare foarte generală pot fi grupate astfel: (1) carcinoame, (2) sarcoame, (3) limfoame, (4) leucemii. Ce cauzează apariţia unui cancer? Declanşarea unui anumit tip de cancer are multiple cauze, iar până la manifestarea clinică sunt necesari câţiva ani sau câteva decenii. Există dovezi certe că iniţierea şi dezvoltarea unui cancer este rezultatul alterării permanente a unor gene aparţinând unei celule sau unui grup de celule. Acestea pot fi (1) înăscute sau (2) achiziţionate de-a lungul vieţii unui individ. Au fost identificate peste 300 de gene care pot fi implicate în procesul de dezvoltare a tumorilor maligne. O alterare la nivelul aşa-numitelor gene ale creşterii cunoscute sub numele generic de oncogene, poate semnaliza celulelor să se dividă în afara controlului. Pe de altă parte, o alterare a genelor supresoare de tumori, care în mod normal servesc ca „întrerupători” ai diviziunilor celulare, vor permite celulelor cu ADN-ul alterat să continue să se dividă, mai degrabă decât să permită repararea ADN-ului si eliminarea celulelor afectate. Agenţii cauzatori ai cancerului pot fi în interiorul sau în exteriorul corpului uman. Tot ceea ce este în afara corpului uman şi interacţionează cu organismul, este denumit mediu. Epidemiologii americani consideră că expunerea la o mare varietate de substanţe naturale sau fabricate de oameni din mediul înconjurător, contează pentru cel puţin 2/3 din toate cazurile de cancer din USA. Există variaţii individuale notabile în ceea ce priveşte susceptibilitatea de a achiziţiona alterarea unor gene cu rol determinant în iniţierea şi susţinerea cancerului: indivizii umani diferă în ceea ce priveşte capacitatea eliminării agenţilor cauzatori de cancer sau de reparare a ADN-ului afectat de agenţii cancerigeni. Genele au un rol minor în majoritatea cancerelor. Factorii de mediu sunt determinanţi principali în declanşarea şi dezvoltarea majorităţii cancerelor. Întrucât, MEDIUL ARE O INFLUENŢĂ MAI PUTERNICĂ ASUPRA RISCULUI DE MANIFESTARE A CANCERULUI DECÂT GENELE, rezultă că noi putem scădea riscul de cancer prin controlarea factorilor care ţin de modul nostru de viaţă: (1) dieta, (2) fumatul, (3) infecţiile, (4) reducerea drastică a poluării. Peste 50% din totalul cancerelor dezvoltate în populaţia umană ar putea fi prevenite dacă s-ar adopta un mod de viaţă controlat: (1) excluderea obiceiului de a fuma şi/sau de a consuma alcool, (2) adoptarea unei diete sănătoase, (3) activitate fizică suţinută, (4) menţinerea greutăţii corporale la valorile normale, (5) şederea prelungită în aer curat sub soare în condiţii de siguranţă. Subtituţia unei singure perechi de baze (point mutation) la nivelul genei p53 poate declanşa diverse tipuri de cancer. Astfel, în cancerul pulmonar, fumul de ţigară (considerat a fi vinovat pentru 90% din totalul neoplasmelor pulmonare) este agentul cauzator al substituţiei bazei G cu T; in cancerul colo-rectal, G este substituită de A; în cancerul hepatocelular, aflatoxina determină „G to T mutation”; în cancerul de piele, radiaţiile UV determină substituţia unui tandem de baze: două baze CC sunt înlocuite de TT. Toate tipurile de substituţii caracteristice acestor feluri de cancer au fost reproduse experimental la animale utilizând agenţii cauzatori menţionaţi. Genele Rb şi p53 (tipul sălbatic) au rol de gene supresoare de tumori. Alterarea mutaţională a genei Rb conduce la apariţia retinoblastomului iar alterarea mutaţională a genei p53 este implicată în apariţia cancerului de sân, creier, a unor ţesuturi moi. Transfecţia in vitro a tipului sălbatic al genei Rb a supresat în grade variate creşterea celulelor, iar după transfectarea tipului sălbatic al genei p53 în unele linii de celule neoplazice (colon, vezică urinară, creier, os) care conţineau gena mutantă p53 s-a realizat supresarea creşterii celulare şi/ sau a tumorigenicităţii.
CONTRIBUŢIA UNOR PROCARIOTE MARINE LA CONSUMAREA DIOXIDULUI DE CARBON ŞI A METANULUI Ioan Ardelean Institutul de Biologie Bucuresti, e-mail: ioan.ardelean@ibiol.ro Microorganismele alcătuiesc un grup vast şi eterogen din punct de vedere al morfologiei, structurii, funcţiilor biologice şi al încadrării sistematice, în care sunt incluse procariote şi unele eucariote: algele microscopice, ciupercile microscopice (mucegaiurile şi drojdiile), protozoarele. Procariotele sunt cele mai active vieţuitoare implicate în circulaţia elementelor chimice în naturǎ, atât în trecutul geologic al Pǎmântului cât şi în prezent. Soarele este principala sursǎ de energie utilizatǎ direct sau indirect de cǎtre fiinţele vii pentru punerea în circulaţie a elementelor chimice în naturǎ, circulaţie de care depinde, printre altele, şi creşterea şi multiplicarea tuturor organismelor, nu numai a microorganismelor. În cursul acestei circulaţii a diferitelor elemente chimice, diferitele tipuri de microorganisme consumă sau produc dioxid de carbon sau metan, gaze care în atmosfera Pământului absorb energie calorică, contribuind astfel la tendinţa de încălzire globală ce caracterizează schimbările climatice. Consecinţele schimbărilor climatice sunt atât de profunde încât comunitatea ştiinţifică are obligaţia să participe la elaborarea politicilor de reducere a emisiilor gazelor ce favorizează efectul de seră şi la direcţionarea metanului şi a dioxidului de carbon spre rezervoare naturale altele decât atmosfera terestră, procese în care participă şi microorganisme, procariote în primul rând, marine. Această prelegere este axată pe contribuţia unor procariote marine la consumarea dioxidului de carbon şi a metanului din următoarele motive mai importante: a) Contribuţia procariotelor fotosintetizante marine (Synechococcus şi Prochlorococcus) este esenţială din punct de vedere cantitativ pentru preluarea dioxidului de carbon din atmosferă, contribuţie care a fost anunţată odată cu cuantificarea abundenţei în mediul marin a genului Synechococcus (1977) şi a genului Prochlorococcus (1987) devenind mai bine cunoscută începând cu 1997; b) Descoperirea asociaţiilor sintrofice de procariote capabile de a oxida metanul în condiţii anaerobe realizată la începutul mileniului III, susţine că acest proces este semnificativ cantitativ oxidând 80% din metanul din mediul marin cum este cazul Mării Negre unde s-au descoperit în 2001 adevărate recife microbiene (microbial reefs); c) Microbiologia marină este o ştiinţă a cărei interdisciplinaritate este în plină creştere (tendinţa fiind de a deveni microbiologie oceanografică) aflată în plină dezvoltare pe plan internaţional, care trebuie promovată şi în ţara noastră inclusiv prin activitatea didactică şi de cercetare.
Consumarea dioxidului de carbon Microorganismele fotosintetizante oxigenice din mediul marin (MFOM) sunt alcătuite atât din procariote cât şi din eucariote (alge unicelulare), împreună constituind doar 1% din cantitatea de carbon care la ansamblul planetei noastre se găseşte stocată în organisme fotosintetice (inclusiv cele pluricelulare). Cantitatea totală de carbon ce se găseşte în biomasa organismelor fotosintetizante oxigenice este estimată la aproximativ 600 de miliarde de tone. Deşi constituie doar 1% din această cantitate uriaşă, activitatea microorganismelor fotosintetizante oxigenice este deosebit de intensă contribuind semnificativ la fluxul de materie şi energie pe planeta noastră. O expresie a acestei activităţi intense a MFOM o constituie capacitatea acestora de a avea o contribuţie foarte importantă la ciclul carbonului de exemplu, preluând din atmosferă cantităţi uriaşe de dioxid de carbon pe care le asimilează reductiv în cursul metabolismului lor, contribuind la sinteză masivă de substanţă organică (care va fi disponibilă diferitelor organisme heterotrofe) consumând totodată cantităţi corespunzătoare de dioxid de carbon, gaz care contribuie la procesul de încălzire globală datorită efectului de seră. Contribuţia majoră a MFOM la circuitul carbonului de exemplu a fost stabilită prin cercetări care au fost iniţiate în anul 1997, atunci când NASA a lansat primul său satelit care avea capacitatea de a monitoriza activitatea MFOM la nivelul Oceanului Planetar. În anul următor, 1998 concluziile cercetărilor efectuate de câteva grupuri de specialişti, în corelaţie cu rezultatele obţinute din satelit, au evidenţiat că MFOM incorporează aproximativ 50 de miliarde de tone de carbon anorganic, ceea ce constituie dublul celor mai optimiste estimări anterioare. Acest tip de cercetări în spaţiul cosmic şi in situ au demonstrat un alt fapt surprinzător: contribuţia fotosintetizantelor terestre la fixarea prin fotosinteză oxigenică a dioxidului de carbon este de aproximativ 52 de miliarde de tone, jumătate din cantitatea estimată prin cercetări clasice, fără utilizarea sateliţilor. În concluzie toate aceste cercetări au arătat că MFOM fixează aproximativ aceeaşi cantitate de carbon ca organismele fotosintetice terestre. Totalitatea organismelor fotosintetizante nu poate asimila excesul de dioxid de carbon a cărui concentraţie creşte continuu în atmosfera Pământului, fiind unul dintre factorii principali care contribuie la încălzirea globală, proces care pare să se fi declanşat cu aproximativ 50 de ani în urmă. Acest proces care îngrijorează nu doar omul obişnuit dar şi oamenii de ştiinţă şi pe politicieni ar putea fi contracarat consideră mulţi specialişti (dar nu toţi!) prin punerea în valoare a capacităţii MFOM de a absorbi cantităţi suplimentare de dioxid de carbon prin intensificarea procesului de fotosinteză oxigenică la scară planetară. Conform unor specialişti, în condiţiile actuale, accelerarea fotosintezei oxigenice ar putea fi posibilă prin aprovizionarea de către om a Oceanului Planetar cu acele elemente chimice care, la ora actuală sunt limitante pentru procesul de fotosinteză. Fundamentul teoretic al acestei ipoteze este următorul: a) În mediul marin intensitatea fotosintezei pare să fie limitată de concentraţiile foarte mici de azot, fosfor şi fier din multe bazine marine; b) Estimarea intensităţii fotosintezei ar conduce la sinteza unei cantităti mai mari de biomasă a MFOM; c) Din care se speră o cantitate din ce în ce mai mare va fi direcţionată spre fundul oceanului unde carbonul nu mai contribuie la efectul de seră.
Consumarea metanului Gazul metan este hidrocarbura cu cea mai mare concentraţie găsită în atmosfera Pământului, contribuind semnificativ la balanţa radiativă a planetei noastre deoarece este un gaz care are un puternic efect de seră comparativ cu alte gaze (de exemplu dioxidul de carbon). În mediul marin principala sursă de metan o constituie sedimentele anaerobe care la anumite locaţii conţin depozite uriaşe de metan. Dacă această cantitate uriaşă de metan ar fi mobilizată brusc de la nivelul sedimentelor în atmosferă concentraţia gazului metan ar creşte semnificativ iar schimbările climatice datorate accelerării procesului de încălzire globală ar fi dramatice. Din fericire pentru umanitate acest proces nu are loc şi se estimează că nici nu se va produce deoarece microorganisme specializate realizează oxidarea metanului; se estimează că microorganisme anaerobe oxidează anaerob metanul în proporţie de 98% ceea ce contribuie semnificativ la diminuarea drastică a cantităţii de metan din mediul marin care ajunge în atmosferă. Oxidarea anaerobă a metanului are loc în sedimentele marine realizată de către consorţii de microorganisme arheene şi bacterii sulfat reducătoare. Asocierea fizică şi fiziologică dintre aceste două tipuri de procariote aparţinând Domeniului Archaea şi respectiv Domeniului Bacteria sunt caracterizate prin procesul de sintrofie. Acest proces este deosebit de important şi pentru Marea Neagră. Marea Neagră este cel mai mare bazin marin meromictic care prezintă în mod constant o mare zonă anoxică care se găseşte sub haloclină; în zilele noastre haloclina se găseşte la adâncimi cuprinse în diverse zone ale Mării Negre între 90 şi 170 de metri. În zona oxică a Mării Negre concentraţia metanului este foarte mică fiind cuprinsă între 1 şi 4 nM semnificativ mai mică decât concentraţia metanului în zona anoxică dedesubtul haloclinei, unde concentraţia este de aproximativ 11μM. În partea de N-V a Mării Negre există numeroase izvoare subterane, localizate în subsuprafaţa fundului marin, izvoare puternice care aduc cantităţi mari dar nedeterminate cantitativ de metan în coloana de apă. Coloana de apă a Mării Negre poate fi clasificată într-o porţiune oxică de dimensiuni relativ mici (5,3 x 1016litri) şi o zonă anoxică cu volum mai mare (4,8 x 1017litri) care împreună conţin aproximativ 96 Tg de metan. S-a estimat că în această zonă anoxică de apă se produce anual o cantitate de 62,9 x 1010moli metan. S-a estimat că volumul de metan eliberat din sedimente împreună cu volumul de metan produs prin metanogeneză este foarte mic comparativ cu volumul de metan eliberat în coloana de apă de către izvoarele subterane. În zona anoxică a Mării Negre valoarea medie a oxidării anaerobe a metanului este de 0,03 nM/zi sub chemoclină atingând o valoare de 3,1 nM la adâncimi mai mari. Oxidarea metanului are loc şi în coloana de apă a Mării Negre, de către microorganisme anoxice în zonele lipsite de oxigen molecular şi de către bacterii metilotrofe în zona oxică a Mării Negre. În concluzie, procariotele marine care consumă dioxid de carbon şi metan au o contribuţie cantitativă semnificativă care diminuează parţial tendinţa de creştere a concentraţiei acestor gaze în atmosfera Pământului, existând preocuparea oamenilor de ştiinţă, a unor politicieni şi chiar a unor firme private de dezvoltare a unor programe regionale sau globale de fertilizare a oceanului pentru intensificarea procesului de fotosinteză oxigenică în mări şi oceane. Dezbaterile ştiinţifice, politice şi economice sunt în plină desfăşurare.
Aurelia BrezeanuInstitutul de Biologie Bucuresti, e-mail: aurelia.brezeanu@ibiol.roÎn ultimii ani o atenţie deosebită se acordă studiului stresului oxidativ, dată fiind atât larga sa incidenţă cât şi efectele dramatice produse, asupra organismelor vii. Pentru a inţelege semnificaţia stresului oxidativ şi mecanismele prin care acestea acţionează asupra proceselor de dezvoltare în general şi a plantelor în particular, se impune a da raspuns la urmatoarele intrebări: Ce se inţelege prin stres oxidativ? Care sunt speciile reactive ale oxigenului şi cine le generează? De ce stresul oxidativ poate fi considerat factor perturbator major al proceselor de dezvoltare? 1. Ce se inţelege prin stres oxidativ. Stresul oxidativ poate fi definit ca acea forma de stres prin acţiunea căreia asupra organismelor vii este stimulată formarea speciilor reactive ale oxigenului (SOR), cu efect perturbator sever asupra metabolismului celular, finalizat cu instalarea colapsului intregului organism. Factorii mediului înconjurător care determină stres oxidativ sunt: poluarea aerului datorită creşterii cantităţii de ozon sau dioxid de sulf; anumite herbicide, de tipul paraquat; metalele grele; secetă; arşită; temperaturi suboptimale; lumină UV; intensităţi foarte ridicate ale luminii care stimulează procesele de fotoinhibiţie; infecţia cu patogeni în decursul senescenţei. Aşadar, stresul oxidativ constituie un element comun diferitelor forme de stres. El reprezintă un mecanism esenţial al dialogului între diferite compartimente celulare, induce modificări ale metabolismului şi proceselor de dezvoltare care cel mai adesea sunt atribuite alterării paternului expresiei genice. 2. Care sunt speciile reactive de oxigen şi cine le generează? Speciile reactive ale oxigenului (SOR) sunt molecule produse în urma unor reacţii redox, în decursul reducerii incomplete a oxigenului sau a oxidării apei prin lanţurile transportoare de electroni, mitocondriale sau cloroplastice. De cele mai multe ori SOR nu acţionează izolat. De pildă, formarea oxigenului singlet (1O2) ca efect al radiaţiilor UV, a fenomenului de fotoinhibiţie a reacţiilor fotosistemului II de transfer de electroni de la nivelul cloroplastului stimulează producerea de alte specii reactive ale oxigenului precum peroxidul de hidrogen (H2O2), anionul superoxid (O2- -), radicalul hidroxil (HO-) şi perhidroxil (O2H.). SOR sunt generate şi în condiţii normale de mediu ca urmare a activităţilor celulare precum fotorespiraţia, b oxidarea acizilor graşi, dar nivelul acestora poate creşte substanţial sub influenţa condiţiilor de stres induse de factorii de mediu. 3. De ce stresul oxidativ poate fi considerat factor perturbator major al metabolismului celular şi al proceselor de dezvoltare? În urma îmbogăţirii atmosferei cu oxigen molecular (O2) cu aproape doua bilioane de ani în urmă, graţie evoluţiei organismelor fotosintetizatoare, speciile reactive ale oxigenului (SOR) au reprezentat un companion incomod al metabolismului aerobic. În contrast cu oxigenul molecular, aceste forme parţial reduse sau derivaţi activaţi ai acestuia (1O2, O2- , H2O2 si OH -) sunt puternic reactive şi toxice, din care cauză induc degradări oxidative severe ale celulelor. În consecinţă evoluţia tuturor organismelor aerobe a devenit dependentă de dezvoltarea unor mecanisme defensive, de apărare, antioxidante, mecanisme “scavenger” eficiente, proprii, menite să elimine SOR. Sistemele antioxidante de apărare includ componente nonenzimatice precum ascorbatul, glutationul redus (GSH), a- tocoferol, carotenoizii si enzimatice: ascorbat peroxidaza, catalaza, dehidroascorbat reductaza, monodehidroascorbat reductaza, superoxiddismutza. Studii de biologie celulara şi moleculară, au permis evidenţierea rolului SOR în controlul şi reglarea unor procese biologice importante precum, moartea celulară programată, reglarea unor procese de semnalizare hormonală ca raspuns la stres indus de atacul patogenilor şi în desfăşurarea unor procese de dezvoltare, precum lignificarea. Recent s-a subliniat rolul dual al SOR în biologia plantelor şi anume pe de o parte bioproduşi toxici ai metabolismului aerobic, iar pe de altă parte reglatori cheie ai unor paternuri defensive (de apărare) la patogeni, ca şi a unor procese de dezvoltare. Ţintele celulare ale SOR sunt: proteinele, ADN, ARN şi lipidele prin a caror oxidare sunt induşi radicali liberi cu efecte toxice severe. Faţă de acţiunea factorilor de stres celular are numai două opţiuni: adaptarea sau moartea (prin necroză sau apoptoză). Termenul adaptarea presupune diferite reacţii celulare care activează mecanismele de rezistenţă şi reparatorii, permiţând integrarea proceselor determinate de stres în funcţionarea normală şi în diferenţierea unor celule. În ce priveste moartea celulară SOR joacă un rol în reglarea atât a necrozei (N) cât şi a apoptozei (MCP). Necroza apare atunci când sub acţiunea unor factori agresivi, în cazul de faţă SOR, celula nu işi mai poate menţine homeostazia ionică şi implicit homeostazia metabolică. Situl major al alterărilor celulare îl reprezintă membrana plasmatică. Apar, depolarizări ale plasmalemei, creşterea concentraţiei calciului citosolic, peroxidarea lipidelor, degradarea tonoplastului şi eliberarea enzimelor lizozomale, vacuolare în spaţiul intracelular, ce conduce la modificări în conformaţia şi funcţia plastidelor şi mitocondriilor. Apoptoza reprezintă o altă formă de rezistenţă la stresul oxidativ la plante, este un proces încă puţin cunoscut în ce priveşte mecanismele moleculare care o guvernează. S-a demonstrat recent la Arabidopsis existenţa unei noi familii de caspaze (cistein proteaze) homoloage la plante, fungi şi protozoare denumite “metacaspaze” cu rol în realizarea fazei de “execuţie” a MCP. Ca şi la animale, la plante mitocondriile joacă un rol important în instalarea MCP intrucât pot să integreze semnale de stres oxidativ care să grabească acest proces. Stresul oxidativ induce formarea la nivelul anvelopei mitocondriale a aşa numiţilor pori de tranziţie (PT) ce facilitează transportul activ de ioni în interiorul matricei mitocondriale, urmat de eliberarea citocromului c; apar perturbari ale transportului ionic transmembranar, creşterea concentraţiei calciului citosolic, ce conduc la activarea “metacaspazelor” şi declanşarea unor dereglări severe la nivel nuclear (clivarea proteinelor citoscheletului nuclear, fragmentări nucleare finalizate cu colaps celular). Paleta alterărilor intracelulare datorată stresului oxidativ creşte pe masura îmbătrânirii organismului. Se postulează că aceasta constituie un factor cauzal al senescenţei. Unul dintre cei mai intens studiaţi factori care induc stres oxidativ este expunerea la concentraţii ridicate de ozon, rezultat al activităţilor umane, motiv pentru care constituie un sistem model pentru demonstrarea modului în care SOR induc alterări oxidative severe la nivelul biomembranelor. Efectul negativ al ozonului asupra plantelor include: descreşterea ratei fotosintezei, degradarea frunzelor, reducerea creşterii tulpinii şi rădăcinii, accelerarea senescenţei şi reducerea producţiei. Mecanismele implicate în producerea alterărilor oxidative induse de ozon (toxicitatea ozonului) nu sunt complet inţelese. Se presupune că pătrunderea sa în spaţiul periplasmic poate fi diminuată de gradul de închidere a stomatelor. Cele mai multe alterări sunt reprezentate de degradarea oxidativă a lipidelor şi proteinelor membranei plasmatice şi producerea de radicali liberi, oxiradicali, sau alţi intermediari reactivi. Ozonul poate reacţiona cu etena şi alte alkene în fluidul apoplastic pentru a forma HO., O2.-, H2O2 şi prin alterarea plasmalemei apar degradări la nivelul permeabilităţii membranare, a transportului de ioni, se inhibă activitatea pompelor H+, are loc colapsul potenţialului de membrană, creşterea concentraţiei ionilor de Ca 2+ captaţi din apoplast. Afectarea ciToplasmei datorată formării radicalilor liberi în interiorul celulei, induce numeroase perturbări în structura fină a organismelor celulare majore, cele mai evidente fiind alterările de la nivelul cloroplastelor şi mitocondriilor. Cu toate ca s-au inregistrat paşi importanţi privind înţelegerea răspunsului plantelor la stresul oxidativ, nu au fost elucidate încă mecanismele implicate. Se presupune că pe măsura progreselor ce se vor înregistra în studiul genomului vegetal, anumite gene asociate cu cascadele de transducţie a semnalelor cunoscute la animale, incluzând receptorii de membrană kinazici şi peptidohormonii, ar putea acţiona şi la plante.
IMPACTUL ANTROPIC ASUPRA ECOSISTEMELOR ACVATICE - EFECTE SI FEED-BACK Cristina Sandu Institutul de Biologie Bucureşti, email: sanducri@yahoo.com Explozia demografică înregistrată în ultimele decenii a dus la degradarea accentuată a mediului ambiant, determinând implicit şi declinul biodiversităţii. Dublarea populaţiei globului în mai puţin de 50 de ani (de la 3 miliarde în 1959 la 6,7 miliarde în 2008, conform US Census Bureau) a avut ca efect modificarea accelerată a utilizării terenurilor (ca urmare a defrişărilor masive, asanării zonelor umede, etc), agricultura şi industrializarea intensivă, poluarea cu nutrienţi sau diferite substanţe xenobiotice, amenajări hidrotehnice (baraje, îndiguiri, amenajarea râurilor pentru navigaţie) şi indirect, încălzirea globală datorită creşterii emisiilor de gaze cu efect de seră. Ca urmare a sporirii populaţiei umane a crescut nevoia de apă pentru consum casnic, agricol sau industrial, ceea ce a făcut ca rezervele de apă dulce sa fie supuse unei degradări accelerate (Postel, 2000). Poluarea chimică afectează grav biocenozele acvatice: deversarea apelor uzate fără un tratament corespunzător, utilizarea unor tehnologii învechite, utilizarea îngraşămintelor şi a pesticidelor în agricultură sunt considerate sursele majore de contaminare (ICPDR, 2005). La nivel mondial se utilizează anual cca 300 milioane tone compuşi chimici, din care mare parte ajung în ecosistemele acvatice, uneori cu consecinţe dramatice (ex. accidentul ecologic de la Baia Mare, 2000). Programele actuale de monitoring sunt orientate cu precădere către determinarea concentraţiei poluanţilor; pentru surprinderea efectelor ecologice ale poluării asupra ecosistemelor acvatice au fost elaborate metode noi, care sa permită evaluarea efectelor sub-letale: biomarkerii moleculari. Un proiect pilot întreprins de colectivul de Ecologie acvatică al Institutului de Biologie Bucureşti sub coordonarea Universităţii Tübingen (Germania) a evidenţiat contaminarea ridicată a unui sector din râul Mureş, deşi parametrii monitorizaţi determinau încadrarea apei în categoria II de calitate (Köhler et al. 2007, Sandu et al. 2008), dovedind necesitatea introducerii unor analize mai sensibile în actualul program de monitoring. Deşi în concentraţii reduse nutrienţii (N, P) au un rol benefic în stimularea producţiei primare, în cantităţi ridicate pot determina eutrofizarea ecosistemelor acvatice, cu efecte negative asupra biodiversităţii. Spre exemplu, cantităţile ridicate de nutrienţi transportate de fluviul Dunărea în perioada anilor `70 au determinat eutrofizarea accelerată a Deltei Dunării după 1980, antrenând o scădere drastică a biodiversităţii de-a lungul întregului lanţ trofic (Brezeanu et al 1991, Vădineanu et al, 2001). Construcţiile hidrotehnice, care au avut ca efect fragmentarea habitatelor, pierderea zonelor umede adiacente, deteriorarea deltelor şi estuarelor, reprezintă una din principalele cauze ale declinului ihtiofaunei: conform World Conservation Union, 20 % dintre speciile de peşti de apă dulce sunt în pericol de dispariţie sau au dispărut deja (Richter & Postel, 2004). Un exemplu dramatic în Bazinul Dunării este declinul sturionilor: din cele 6 specii native, una este considerată dispărută (Acipenser sturio), celelalte fiind clasificate ca vulnerabile sau periclitate (IUCN 2004, în Bloesch et al, 2006). Includerea Coridorului Verde al Dunării în Coridorul VII de transport pan-european şi transformarea acestuia în cale de navigaţie maritimă, va avea ca efect accentuarea impactului negativ asupra biodiversităţii. Pe lângă aceste efecte directe asupra ecosistemelor acvatice, în ultimele decenii au început să fie resimţite efectele indirecte ale încălzirii globale: Topirea gheţarilor şi a calotelor glaciare, creşterea temperaturii şi a nivelului mărilor şi oceanelor, schimbarea regimului precipitaţiilor şi creşterea riscului producerii unor fenomene meteo extreme (secete, inundaţii), reducerea rezervelor de apă dulce, etc (IPCC 2008). Modificarea regimului precipitaţiilor a avut efecte negative asupra debitului Dunării, acesta fiind în scădere după 1960 (Michaylova, 2004), fapt ce a afectat şi ecosistemele lacustre din Delta Dunării (Wehrli et al, 2008). Degradarea mediului şi scăderea biodiversităţii nu înseamnă însă numai dispariţia unor specii ci şi pierderea unor bunuri şi servicii vitale existenţei umane. Apa este un element indispensabil vieţii, iar ecosistemele acvatice au un rol esenţial în purificarea apei, circuitul nutrienţilor, reglarea climatului şi compoziţiei atmosferice, temperarea viiturilor şi secetelor, turism şi recreere (Turner et al, 2007). De aceea este important sa fie găsite soluţii viabile pentru conservarea acestor ecosisteme şi pentru atenuarea impactului negativ al dezvoltării umane. Bibliografie BLOESCH, J., JONES, T., REINARTZ, R. & STRIEBEL, B. (2006): An Action Plan for the conservation of sturgeons (Acipenseridae) in the Danube River Basin. ÖWAW 58/5-6: 81-88. BREZEANU, G., ZINEVICI, V., NICOLESCU, D., NICOLESCU, N. (1991): Die Strkturellfunctionelle Evolution der Biozonosen aus den Seen des Donaudeltas unter den einfluss der Eutrophiesierung, 29 Arbeits der IAD, Kiev, p. 275-281. ICPDR (2005): The Danube River Basin District Part A - Basin-wide overview. WFD Roof Report 2004, Vienna. [IPCC] INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE (2008): Climate change and water. IPCC Technical Report VI, 214 p. KÖHLER, H-R., SANDU, C., SCHEIL, V., NAGY-PETRICĂ, E.M., SEGNER, H., TELCEAN, I., STAN, G. & TRIEBSKORN, R. (2007): Monitoring Pollution in River Mureş, Romania, Part III: Biochemical Effect Markers in Fish and Integrative Reflection. Environmental Monitoring and Assessment, 127: 47-54. MICHAYLOVA, V. N. (2004): Hydrology of Danube Delta, Moscow (in Russian). POSTEL,S.L. (2000): Entering an era of water scarcity: The challenges ahead. Ecological application, 10:941-948. RICHTER, B., POSTEL,S.L. (2004): Saving Earth’s rivers. Issues in Science and Technology, 31-36. SANDU, C., FARKAS, A., MUSA-IACOB, R., IONICA, D., PARPALA, L., ZINEVICI, V., DOBRE, D., RADU, M., PRESING, M., CASPER, H., BURUIANA, V., WEGMANN, K., STAN, G., BLOESCH, J., TRIEBSKORN, R., KÖHLER, H.-R. (2008): Monitoring Pollution in River Mureş, Romania, Part I: The limitation of traditional methods and community response. Archiv fur Hydrobiologie, Suppl. Large Rivers, 18 (1-2): 91-106. TURNER, W., BRANDON, K., BROOKS, T., COSTANZA, R., DaFONSECA, G.A.B, PORTELA, R. (2007): Global Conservation of Biodiversity and Ecosystem Services. BioScience, Vol. 57, 10:868-873 VADINEANU. A., CRISTOFOR, S., IORDACHE, V. (2001): Lower Danube River System biodiversity changes. In: Gopal, B., Junk, W.J., Davis, J. (eds.) Biodiversity in Wetlands: Assessment, Function and Conservation. Backhuys Publishers, p. 29-63. WEHRLI, B., SANDU, C., LYASHENKO, A., AFANASIEV, S. (eds.) (2008): Impact of environmental changes on aquatic ecosystems of Danube Delta (ECAQUDAN). SCOPES Project report, in preparation.
BIOINDICATORII ŞI SEMNIFICAŢIA SCALEI SPAŢIO-TEMPORALE ÎN ASPECTELE LEGATE DE SCHIMBĂRILE CLIMATICE Dorina Purice Institutul de Biologie Bucureşti, email: dorina.purice@ibiol.ro; dorina_purice@yahoo.com
În ultimii 20 de ani unele dintre problemele globale majore privind mediul înconjurător s-au identificat a fi reducerea drastică a resurselor neregenerabile, deteriorarea severă a capitalului natural şi schimbările climatice. Cauzele determinante ale acestor fenomene se manifestă sinergic şi transfrontieră. În cercetările ecologice din ultimele decenii se utilizează din ce în ce mai mult bioindicatorii, ca instrumente de diagnoză privind multitudinea de forme de impact antropic asupra calităţii sistemelor ecologice. Se utilizează această manieră de abordare, împreună cu alte direcţii de cercetare, datorită nevoii de cuantificare cât mai rapidă a unor probleme complexe. Succesul unor asemenea evaluări depinde de stabilirea iniţială corectă a termenilor de studiu, în special a scalei spaţio-temporale. Una dintre direcţiile urmate în studiile ce vizează speciile bioindicatori este diversitatea (la diferite scale) în arealele vizate. Loreau et al. (2001) arătau că tipurile de impact uman asupra mediului înconjurător la scale locale până la globale cauzează nu numai un declin al diversităţii, ci şi schimburi previzibile funcţionale pe măsură ce anumite grupuri de specii cu caracteristici particulare sunt înlocuite de alte grupuri de specii cu caracteristici diferite. Schimbările climatice conduc la o pierdere globală a speciilor pe măsură ce condiţiile abiotice încep să depăşească limitele de tolereanţă ale speciilor. Bohác et al. (1991) menţionează faptul că populaţiile şi comunităţile de nevertebrate sunt utilizate în principal pentru investigaţii la scală locală. Aceste grupe au dimensiuni mici ale corpului şi o mai mică tendinţă de migraţie, şi sunt de aceea adecvate pentru indicarea factorilor de mediu la nivel local (aplicaţii neadecvate ale fertilizatorilor şi pesticidelor, management inadecvat al complexelor de ecosisteme etc.). Este deja bine ştiut că procesele ecologice nu sunt independente de spaţiul în care se petrec şi că ele sunt “articulate”, conectate spaţial. Astfel, în timp ce modelele asociative pot fi utilizate în “universul” timpului şi spaţiului din care sunt colectate datele, nu există nicio cale de evaluare a extinderii la care ele pot fi utilizate în alte condiţii în care relaţiile şi asociaţiile nu pot fi aplicate (de la Peña et al, 2003). În ideea de a produce modele generalizabile pentru predicţiile în care sunt utilizate de exemplu carabidele (binecunoscute şu utilizate ca bioindicatori), avem nevoie să luăm în considerare procesele ecologice evidente precum şi spaţiul în care se petrec. Unei specii i se poate atribui un model la nivel de individ (indivizi), populaţie sau metapopulaţie. În mod clar, încercarea de a modela de exemplu efectele schimbării utilizării terenului asupra individului, este nerealistă. La scala spaţială mai mare, organismele pot fi considerate a fi membri unor populaţii distincte. În interiorul populaţiilor, indivizii interacţionează într-un anume fel, de obicei prin activitate reproductivă sau prin competiţia pentru resurse. La scală spaţială încă şi mai mare, populaţiile pot fi abordate ca grupuri de populaţii (părţi ale unei metapopulaţii) care se întâlnesc în unităţi semi-izolate ale habitatului. Populaţiile individuale interacţionează în metapopulaţie destul de liber, prin dispersie. Din moment ce modelele metapopulaţiei analizează schimbările în distribuţia populaţiilor în funcţie de probabilităţile exticnţiei şi colonizării peticelor de habitat, aceste modele sunt de o oarecare utilitate practică în cercetarea la scala complexului de ecosisteme. Hunter (2002) spunea: “Dacă spaţiul este frontiera finală a teoriei ecologice (Kareiva, 1994), atunci fragmentarea spaţiului este motorul “warp” care conduce cercetarea în ecologia spaţială”. Raportul marginii habitatului către interior, izolarea fragmentelor de habitat, zona peticelor, calitatea şi diversitatea peticelor precum şi microclimatul, toate contribuie la determinarea abundenţei şi bogăţiei insectelor în complexele de ecosisteme. Dată fiind importanţa mişcării în ecologia spaţială a insectelor, nu ar trebui să fie o surpriză faptul că mărimea şi dispunerea fizică a peticelor de habitat asupra complexelor de ecosisteme joacă un rol fundamental în determinarea abundenţei şi diversităţii faunei de insecte. Hunter (2002) afirmă că realizarea faptului că lungimea lanţului trofic şi a structurii trofice sunt aşa de vulnerabile la fragmentare este cea mai importantă lecţie pe care am învăţat-o din studiul insectelor, la nivelul complexului de ecosisteme. Răspunsurile diferenţiate ale prădătorilor şi prăzii la structura complexului de ecosisteme par să se menţină atunci când studiile se realizează la scale mai mari. Fragmentele mici şi dispersate de habitat sunt cunoscute că sporesc diversitatea beta în complexele de ecosisteme. Tscharntke et al. (2002) subliniază importanţa fragmentelor mici de habitat pentru anumite specii de insecte şi punctează faptul că cel puţin pentru anumite grupe de fluturi, fragmentele mici favorizează existenţa mai multor specii decât o fac zonele echivalente în petice mai mari. Pentru scopuri de conservare, ei recomandă o combinaţie de petice largi cu fragmente mici răspândite pentru a maximaliza diversitatea beta. Pentru stabilirea efectelor schimbărilor climatice la scală spaţio-temporală mai mare, se renunţă la utilizarea speciilor bioindicatoare şi se utilizează indicatori ecologici de tipul ecosistemelor şi al complexelor de ecosisteme (de ex. statutul şi dinamica pădurilor, a zonelor umede etc. pe o anumită perioadă de timp şi la o anumită scală spaţială). BIBLIOGRAFIE SELECTIVĂ
Liliana Vasiliu-Oromulu Institutul de Biologie Bucureşti, email: liliana_oromulu@yahoo.com
În ce fel este influenţatǎ biodiversitatea de impactul factorilor de mediu? Care sunt cauzele acestor influenţe şi care sunt consecinţele ecologice? În mǎsura în care biodiversitatea va fi modificatǎ şi ecosistemele vor fi modificate, iar produsele şi serviciile ecologice indispensabile dezvoltǎrii durabile vor fi ameninţate. Cum reacţioneazǎ insectele tisanoptere la schimbǎrile factorilor de mediu? Ritmul acestor trenduri depinde de condiţiile de habitat, de management şi de biologia reproductivǎ a thripşilor. Tisanopterele sunt insecte pterigote cu dimensiuni reduse, de la 0,5 la 1-2 mm în zonele temperate şi până la 15 mm în cele tropicale. Sunt răspândite pe toate continentele, de la nivelul mării până la cele mai mari altitudini; majoritatea sunt fitofage, unele zoofage, se dezvoltă pe diferite substraturi, unele specii sunt dăunătoare plantelor cultivate în natură şi în sere, altele sunt utile, polenizatoare. Impactul factorilor de mediu a fost relevat într-un studiu de monitoring efectuat în Masivul Gârbova, între 1967 - 1998, în 6 situri de pajişti secundare situate altitudinal între 800 – 1500 m. Analiza temporală a structurii taxonomice a tisanopterelor relevă două perioade distincte în dinamica acesteia: prima este caracteristică perioadei 1967-1982, iar cea de a doua, perioadei 1982-1998. În anul 1982, tisanopterele au suferit o reducere masivă a spectrului taxonomic, de pânǎ la 57,10%, aceasta datorită impactului modificărilor ambientale (începerea unei perioade de secetă accentuată de circa 12 ani şi o creştere evidentǎ a temperaturilor). Diversitatea tisanopterelor relevă o revenire la complexitatea specifică anterioară abia din anul 1995. Bogǎţia de specii de thripşi din pajiştile altitudinal similare din Masivul Gârbova şi Masivul Bucegi, este comparativ de 2,4 ori mai mare la 1163 m în Masivul Gârbova şi respectiv de 1,6 ori la 1500 m, fapt datorat factorilor de mediu: intensitatea vântului diminuatǎ în Masivul Gârbova şi indicelui DeMartonne de 40 în Masivul Gârbova şi respectiv 50 în Masivul Bucegi. În suprafeţele experimentale de pe Muntele Blana, din Masivul Bucegi, supuse renaturalizǎrii prin însǎmânţǎri cu diferite poacee, diversitatea specificǎ a thripşilor a crescut treptat în primii doi ani dar, datoritǎ temperaturilor foarte scǎzute, zǎpezilor abundente şi vânturilor puternice în al treilea an, tisanopterele au fost complet eliminate. În studiul tisanopterelor din parcurile bucureştene, impactul factorilor de mediu a fost relevat atât la nivel taxonomic cât şi populaţional. Dacǎ în primul an, 2006, spectrul specific a fost constituit din 14 specii de thripşi şi o densitate numerică de 3140 ind/m2, în 2007 datoritǎ temperaturilor excesive din timpul verii prelungite, s-a remarcat înlocuirea speciilor mezofile cu cele termofile, biodiversitatea fiind mai amplǎ, de 35 specii dar cu densitate numericǎ redusǎ, de 2042 ind/m2, caracteristice fiind 3 specii bioindicatori sensibili ai poluǎrii atmosferei. Pe haldele din Masivul Retezat, echilibrul populaţional al tisanopterelor a fost complet perturbat de ploile diluviene din 1999, tendinţa de refacere a diversitǎţii acestor insecte a început abia dupǎ doi ani, de la acest impact de mediu. Speciile de tisanoptere native din America sau Australia, pǎtrunse în Europa au devenit specii invazive, având în serele din Europa pânǎ la 8 – 12 generaţii/an, datoritǎ factorilor de mediu favorabili, creaţi artificial în aceste spaţii închise. În ecosistemele forestiere speciile de thripşi pot fi dǎunǎtoare prin atacul acestor insecte, coroborat cu impactul factorilor de mediu. În agroecosisteme, speciile de thripşi pot produce pagube economice în funcţie de condiţiile de mediu, de efectul sinergic al atacului insectei şi al virusurilor (din Fam Bunyaviridae) transmise plantelor. Caracteristicile ecologice ale tisanopterelor, biologia lor, lista roşie, toate sunt influenţate şi de impactul factorilor de mediu.
IMPACTUL POTENŢIAL AL SCHIMBĂRILOR CLIMATICE ASUPRA ECOSISTEMELOR ALPINE SORIN ŞTEFĂNUŢ Institutul de Biologie Bucureşti, e-mail: sorin.stefanut@ibiol.ro Schimbările climatice din ultima sută de ani au afectat speciile şi ecosistemele, în special cele alpine. Ecosistemele alpine sunt în mod particular foarte sensibile la schimbarile climatice, aşa cum au relevat cercetările recente efectuate în ultimii ani în Alpi, Europa (Holzinger & al. 2008; Parolo & Rossi 2008). Studiile efectuate în aceaste zone au arătat că în ultima jumătate de secol, temperaturile medii anuale din timpul verii au crescut cu 1,6 ºC şi cu 1,1 ºC temperaturile medii anuale din timpul iernii. În aceiaşi perioadă, izotermele de - 2 ºC şi -1 ºC au urcat cu 240 m în altitudine. Investigaţiile efectuate în ultimul deceniu în Alpi au dus la concluzia că, modificări importante în compoziţia vegetaţiei alpine au avut loc în ultima sută de ani şi cu precădere în ultimii 50 de ani (Grabherr & al. 1994; Camenish 2002; Walther & al. 2004). Comparând datele istorice (1954-1958) cu rezultate recente (2003-2005) a fost identificată o creştere a numărului de specii din zona alpină prin urcarea în altitudine a speciilor din etajul subalpin. De asemenea, 52 de specii alpine au fost identificate la o altitudine cu 30-430 m mai sus decât limita lor din anii 1950, ceea ce corespunde cu o rată de migrare de 23,9 m/decadă. Cercetările din Alpi sunt încă la început şi efectuate doar pe plante superioare şi anumite tipuri de asociaţii alpine. Zonele alpine oferă bune oportunităţi pentru studiul influenţei schimbărilor climatice asupra plantelor superioare, briofitelor, macromicetelor si vegetaţiei, deoarce acestea sunt guvernate de factori abiotici precum temperatura, precipitaţiile şi stratul de zapadă, direct influenţaţi de încălzirea globală. Studiile nostre efectuate pe diferite specii alpine şi subalpine de plante, în perioada 1997-2008, au dus la concluzia că specii subalpin-alpine au urcat 250 de m în altitudine, retragându-se în zona alpină. Spre exemplu, hepatica Bucegia romanica, descrisă pentru prima oară de Radian S.Ş. de pe Valea Cerbului, Munţii Bucegi, a fost colectată în anul 1899 de la o altitudine de 1500 m. De asemenea, Ştefureac T. descoperă în anul 1946 aceasta specie în Munţii Piatra Craiului începând de la o altitudine de 1300 m, având un optim de creştere la altitudini de 1600-1850 m, fiind din ce în ce mai rară spre 2300 m. În urma cercetărilor noastre efectuate în masivele Bucegi, Piatra Craiului, Fagaraş şi Rodnei, specia nu a mai fost regăsită sub altitudinea de 1750 m, fiind însă identificată frecvent la altitudini de 2000-2300 m.
IMPACTUL SCHIMBǍRILOR CLIMATICE ASUPRA INSECTELOR Sanda Maican Institutul de Biologie Bucureşti, email: sanda.maican@ibiol.ro
Se afirmă că trăim în era insectelor, deoarece: - se estimează că în natură ar exista între 50 şi 70 milioane de specii de insecte dintre care doar circa 1,5 milioane sunt descrise; - din totalul de 1,7 milioane specii de organisme cunoscute, insectele reprezintă 55%, comparativ cu plantele cu flori (14%), celelalte nevertebrate (16%), vertebrate (3 %) etc.; - omul consumă ceea ce rămâne în urma atacului insectelor, pierderile de recoltă, care pot merge până la compromiterea completă a culturii, fiind cuprinse între 25%-50% din producţia potenţială.
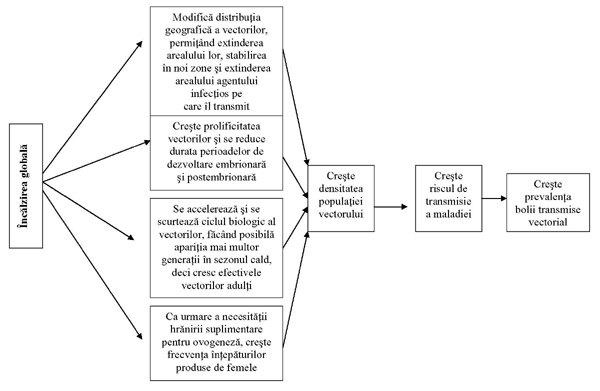
Modificările climatice majore constau din: creşterea temperaturii medii a oceanelor şi atmosferei, modificarea cantităţii şi regimului precipitaţiilor, modificarea cantităţii evaporaţiei. Efectele creşterii temperaturii globale medii: creşterea nivelului oceanului planetar, modificarea circuitului global al apei, inundarea unor mari suprafeţe de uscat, modificarea distribuţiei şi compoziţiei florei şi faunei. Consecinţe ale creşterii nivelului planetar: inundarea terenurilor joase, creşterea frecvenţei inundaţiilor temporare, inundarea plajelor, eroziunea dunelor, salinizarea apei în estuarele râurilor, inundarea zonelor umede situate de-a lungul râurilor; influenţe directe asupra distribuţiei şi diversităţii florei şi faunei, inclusiv a grupului dominant (insectele). Modificările climatice antrenează creşterea frecvenţei şi intensităţii fenomenelor meteorologice extreme, ca seceta şi inundaţiile (cu afectarea ciclului de dezvoltare al insectelor acvatice şi faunei din sol). Se presupune că, datorită vitezei cu care se produc modificările climatice, speciile vegetale nu vor putea fi capabile să se disperseze suficient de repede pentru a supravieţui. Orice modificare în compoziţia specifică şi productivitatea plantelor (suportul vieţii de pe Terra) antrenează modificări în cascadă la nivelul consumatorilor de diferite ordine, afectând în principal consumatorii primari (insectele fitofage). Efectele creşterii temperaturii (factor important care influenţează dezvoltarea şi reproducerea insectelor în natură) asupra insectelor: extinderea arealului unor specii, reducerea biodiversităţii, mărirea biodiversităţii, uniformizarea biodiversităţii la nivel global, reducerea duratei ciclurilor de viaţă, scurtarea perioadei sau eludarea diapauzei hibernale. Modificări mici ale temperaturii, precipitaţiilor şi evaporării pot determina diferenţe regionale mari, inducând modificări în distribuţia florei şi faunei. Se estimează că o creştere cu 10C a temperaturii medii anuale în Europa va determina împingerea arealului unor specii către nord, cu sute de kilometri. Evaluarea toleranţei la temperatură a demonstrat faptul că speciile de insecte din zonele temperate se adaptează mult mai uşor la schimbările climatice, în timp ce pentru cele din zonele tropicale, creşterile de temperatură afectează însăşi supravieţuirea lor. La creşteri nesemnificative de temperatură, rata reproducerii la insectele din regiunile tropicale tinde să se menţină în limite normale; atunci când valorile termice cresc cu 2-40C, rata de reproducere scade accentuat, punând în pericol existenţa speciilor respective. Diapauză - hibernare, estivaţie (strategii de adaptare ale insectelor la condiţii fluctuante de temperatură, definiţie, exemple). Efectele încălzirii globale asupra polenizǎrii plantelor. Scăderea abundenţei sau dispariţia insectelor polenizatoare poate avea consecinţe dramatice în special asupra plantelor care depind de un singur polenizator. Încălzirea globală afectează polenizarea pe care o asigură insectele pentru anumite culturi, reducând producţia acestora. Efectele încălzirii globale asupra sǎnǎtǎţii umane. · creşterea efectivelor unor specii de insecte vectoare pentru boli infecţioase şi parazitare are ca rezultat creşterea numărului acestor boli; · reemergenţa unor boli eradicate; · extinderea arealului unor specii de insecte vectoare pentru boli infecţioase şi parazitare; · extinderea arealului unor specii de patogeni şi paraziţi (virusuri, bacterii, ciuperci, protozoare, helminţi, insecte) care pot produce boli grave. Exemple (malaria, filariozele limfatice, leishmanioze).
INFLUENŢA FACTORILOR DE MEDIU ASUPRA BIODIVERSITĂŢII FAUNEI DE SOL DIN ECOSISTEMELE FORESTIERE Viorica Honciuc Institutul de Biologie Bucureşti, email: viorica.honciuc@ibiol.ro, v_honciuc@yahoo.com În cadrul sistemului biologic superior, biosfera, solul constituie alături de ocean, unul din rezervoarele şi uzinele vieţii de pe planeta noastră. La nivelul lui se desfăsoară cu intensitate maximă procesele prin care ceea ce este mineral devine organic, iar deşeurile organice ale vieţii sunt mineralizate şi redevin apte să intre în circulaţie biologică.. Ca formaţie naturală, solul prezintă un caracter unic : acela de a fi totodată un produs biologic şi un habitat natural, deosebit de complex, animat de o populaţie densă, extrem de variată. Aceste două aspecte, care nu pot fi separate decât în mod conveţional, sunt atât de intim legate, încât dincolo de discontinuitatea şi heterogenitatea aparentă, morfologică, a componentelor sale, conferă solului o omogenitate funcţională care a permis compararea lui cu un ‘’organism viu ‘’. Încă de la începutul secolului trecut se menţiona, că solul, fiind o formaţiune naturală înzestrată cu materie organică proprie (humusul) şi sediul unor procese fiziologice şi biochimice complexe aflate în interacţiune, prezintă numeroase analogii cu organismele vii. Solul « digeră » materiile organice încorporate « asimilează », sintetizând substanţe organice proprii, «respiră », consumând O2 şi elimină CO2. În realitate, solul este – în cadrul ecosistemului - un sistem biologic deschis şi animat de imensitatea organismelor şi microorganismelor ce-l populează, de aceea extrem de dinamic, aflat într-un schimb continuu de energie şi materie cu mediul înconjurător. Rezultatele a numeroase cercetări din ultimile decenii au demonstrat rolul diversităţii animalelor din sol în grăbirea proceselor de humificare şi mineralizare a resturilor de plante, în îmbogătirea particularităţilor structurale ale solului, în creşterea şi îmbogăţirea în substanţe organice etc. În ecosistemele naturale şi în cele create de om, animalele din fauna de sol prin diversitatea lor influienţează descompunerea materiei organice în interacţiunea cu microorganismele. Procesul de descompunere în prezenţa animalelor este de două ori mai rapidă decât în absenţa lor. Cu privire la conceptul « fauna solului » sunt date în literatură multe definiţii dintre care cea mai adecvată pare a fi cea a lui Van der Drift (1955). El consideră ca « fauna solului » include acele animale care îşi petrec în sol întreaga viaţă sau numai o parte din ciclul vital, numai stadiul corespunzător fiind considerat ca aparţinând solului. Pentru marea diversitate de animale care locuiesc în sol s-au adoptat diferite criterii convenţionale de clasificare, cum sunt : dimensiunea corpului (microfauna, mezofauna, macrofauna, megafauna), adaptare la condiţii de umiditate (higrobionte,higrophile, mezophile), de hrana (fitofage, zoofage, necrofage, micofage, saprofage), legătura cu solul ca loc de trai (geobionte, geophile, geoxene) etc. Majoritatea acestor grupe se regăsesc în structura solurilor forestiere, unde participă în mod direct sau indirect la descompunerea materiei vegetale până la humificare. Factorii de mediu ca principali reglatori ai populaţiilor de animale din fauna de sol, acţionează în primul rând asupra solului în ansamblul lui, a tuturor organismelor care stau la baza vieţii solului. În acest sens cei mai importanţi factori sunt: aerul, apa şi temperatura. Aerul din sol prin cantitatea şi compoziţia sa acţionează ca un factor important pentru desfăşurarea proceselor vitale. Unul din gazele cele mai importante, existente în aerul din sol este CO2 , produsul final al descompunerii substanţelor organice. Aerul solurilor de pădure conţine, aproape întotdeauna cantităţi mai mari de CO2 , decât acela al solurilor cultivate. Animalele reacţionează diferit la conţinutul CO2 . După unele observaţii cârtiţa este deosebit de sensibilă la o concentraţie ridicată de CO2 în aerul din sol. Potrivit cercetărilor animalele din sol prezintă diferite grade de sensibile faţă de concentraţii mari de CO2. Pe baza unor experimente s-a stabilit că larvele de elateride sunt mai sensibile decât râmele, iar larvele cărăbuşilor s-au dovedit deosebit de rezistente. În privinţa nematodelor, colembolelor şi acarienilor, este puţin cunoscută influienţa bioxidului de carbon. Influienţa apei asupra vieţii din sol se manifestă cantitativ cât şi calitativ. Apa se află în sol, atât în stare solidă cât şi lichidă sau gazoasă. Numai în ultimile două stări ea are o importanţă bioedafică deosebită influienţând activitatea şi diversitatea faunei de sol. Apa din sol, provine în primul rând din precipitaţii şi numai în mică măsură din apa freatică şi din umiditatea atmosferică. Se poate demonstra dependenţa animalelor din sol faţă de conţinutul în apă al solului. Această relaţie se reflectă cel mai pregnant la ciliate, tardigrade, rotiferi şi nematode care se închistează în condiţii nefavorabile de umiditate, adică îşi suspendă viaţa activă până când mediul înconjurător conţine din nou apă suficientă. Majoritatea animalelor din sol sunt animale de “aer umed”. Acest fapt arată că ele nu-şi pot desfăşura activitatea decât într-un aer al solului care este saturat cu vapori de apă. Se consideră că deosebirile sezoniere calitative şi cantitative ale faunei din sol trebuie atribuite diferenţelor survenite în umiditatea solului. În această situaţie sunt solurile forestiere a căror zone mai profunde, unde regimurile de umiditate sunt mai uniforme, variaţiile sezoniere apar în general mult mai puţin evident. În schimb umiditatea excesivă, provoacă deseori o migrare a animalelor. Astfel se observă din când în când apariţia colembolelor pe suprafaţa zăpezii, sau apariţia frecventă a acestora şi a acarienilor în cazul umezirii intense a unei grămezi de compost forestier. Un alt factor - temperatura solului acţionează asupra activităţii biologice din sol, influienţând astfel humificarea şi mineralizarea substanţelor organice şi alterarea biochimică a celor anorganice din sol şi în acest fel indirect şi fauna de sol. Sursa principală de căldură o constituie iradierea solară. Alte surse de căldură, ca de exemplu căldura internă a planetei şi cantităţile de căldură eliberate în cursul reacţiilor biochimice influienţează numai în foarte mică măsură temperatura solului. O parte din energia radiată de soare este absorbită de suprafaţa solului, iar altă parte este reflectată în atmosferă. Energia solară absorbită încălzeşte solul iar cea reflectată, atmosfera. În funcţie de toleranţa la căldură animalele se împart în 3 categorii: psihrofile (iubitoare de frig), cu o limită inferioară de 0-10, una superioară de -25 oC; aceea a celor mezofile având o temperatură optimă de 25-35 oC şi cea a celor termofile (iubitoare de căldură), cu o temperatură minimă cuprinsă intre 25 si 45 oC şi una maximă de 75-80oC. Între aceste categorii există numeroase tranziţii. Fauna de sol din ecosistemele forestiere aparţine în cea mai mare parte grupei mezofile. Animalele din sol reacţionează în mod specific la modificările de temperatură. Pe lângă aceştii factori analizaţi mai sunt şi alţii, care acţionează indirect asupra diversităţii faunei de sol, fiind mai puţin restrictivi cum sunt: textura şi structura solului, proprietăţile de adsorbţie, reacţie şi potenţial redox ale solului, energia radiantă şi acţiunea complexă a factorilor staţionali.
BIODIVERSITATE, STABILITATE ŞI PRODUCTIVITATE ÎN CONDIŢII ECOLOGICE NATURALE ŞI DE IMPACT ANTROPIC Laura Parpală, Victor Zinevici, Doina Ionică, Mirela Moldoveanu, Larisa Florescu Institutul de Biologie Bucureşti, email: lauraparpala@yahoo.com
A. BIODIVERSITATEA Biodiversitatea, după definiţia Uniunii Internaţionale pentru Conservarea Biodiversităţii, cuprinde toate formele de viaţă, ecosistemele şi procesele ecologice şi ia în considerare ierarhia lor la nivel genetic, specific şi ecosistemic. Biodiversitatatea menţine procesele ecologice la nivel local, regional şi global, generează bunuri şi servicii direct utilizabile sistemului socio-economic. Activitatea umană determină, din ce în ce mai mult, reducerea biodiversităţii, periclitând stabilitatea ecosistemică, cu implicaţii economice, ştiinţifice, estetice şi etice. Biodiversitatea are patru componente ierarhice: diversitatea specifică, diversitatea genetică, diversitatea ecologică (ecosistemică), diversitatea antropică sau etnoculturală. I. DIVERSITATEA SPECIFICĂ Priveşte varietatea speciilor la nivel local (biocenoză), regional (biom) şi global (biosferă). Până în prezent au fost descrise cca. 1,6 milioane specii şi se estimează ca ar fi între 5 şi 10 milioane specii. Numărul de specii descrise şi numărul estimat de specii existente
II. DIVERSITATEA GENETICĂ Rata modificărilor evolutive ale unei populaţii este proporţională cu gradul de diversitate genetică. Genofondul biosferei conţine întreaga informaţie asupra proceselor biologice planetare; diminuarea acestuia limitează capacitatea populaţiilor de a se adapta la modificările mediului şi reduce informaţia biologică potenţial utilă pentru societatea umană. Rolul geneticii ecologice este de păstrare a diversităţii (Wilson şi colab., 2005). III. DIVERSITATEA ECOLOGICĂ (ECOSISTEMICĂ) Distrugerea unor categorii de ecosisteme şi dispariţia rapidă a unor specii, impune necesitatea evaluării rolului speciilor în cadrul ecosistemului în vederea cunoaşterii modului în care diversitatea specifică influenţează structura şi funcţionarea ecosistemelor. În acest sens există mai multe criterii de clasificare a ecosistemelor: · după tipul major de bioTop – sisteme acvatice şi terestre, · după impactul antropic – naturale, seminaturale şi antropizate, · după caracteristicile structurale – abundenţa şi distribuţia speciilor, durată de viaţă, importanţă etc, · după caracteristicile funcţionale – capacitatea productivă a ecosistemelor, de circulaţie a materiei şi energiei de la un nivel trofic la altul, · după natura şi importanţa bunurilor şi serviciilor furnizate sistemului socio-economic, · după capacitatea de refacere sau rezistenţa la modificări = rezilienţa ecosistemelor. IV. DIVERSITATEA ANTROPICĂ SAU ETNOCULTURALĂ Se referă la diversitatea etnică, lingvistică şi culturală a comunităţilor umane. Sistemul socio-economic a evoluat lent multă vreme. Odată cu revoluţia industrială, societatea umană ia din natură resurse şi energie şi returnează deşeuri, ceea ce conduce la reducerea biodiversităţii şi perturbarea stabilităţii sistemului global al Terrei. Toate acestea au condus la elaborarea Programului Naţiunilor Unite pentru Mediu şi Dezvoltare. B. STABILITATEA Stabilitatea reprezintă o stare a ecosistemelor la care se ajunge prin funcţia de autocontrol care, ca şi funcţia de circulaţie a materie şi funcţia energetică, este rezultatul interdependenţei dintre speciile componente, precum şi dintre acestea şi factorii de mediu. Rolul autocontrolului este de a păstra o stare de echilibru între populaţiile componente, de a nu permite oscilaţii numerice prea mari, ceea conduce la stabilitate în structura şi funcţionarea întregului ecosistem (Botnariuc, 1982). Elton (1958) arată că stabilitatea biocenozei este direct proporţională cu creşterea complexităţii reţelei trofice. Reducerea şi/sau înmulţirea excesivă a diversităţii speciilor poate duce la pierderea stabilităţii şi funcţiei ecosistemelor C. PRODUCTIVITATEA Productivitatea reprezintă ”rata” de acumulare a materiei organice sau energiei unei populaţii sau biocenoze, într-o anumită unitate de spaţiu/volum şi timp. În strânsă corelaţie cu bogăţia specifică, productivitatea are valori diferite, eficienţa ei fiind mai mult sau mai puţin utilă pentru om. Impactul antropic datorat, pe de o parte intervenţiei umane, iar pe de altă parte unor cauze naturale (factorii de mediu), este principalul răspunzător al modificării pe termen lung a diversităţii specifice şi pe termen scurt a abundenţei numerice, biomasei şi productivităţii, de regulă în defavoarea sistemului socio-economic. Aceste modificări sunt evidenţiate de noi în ecosistemele lacustre din Delta Dunării, pe o perioadă de 30 ani.
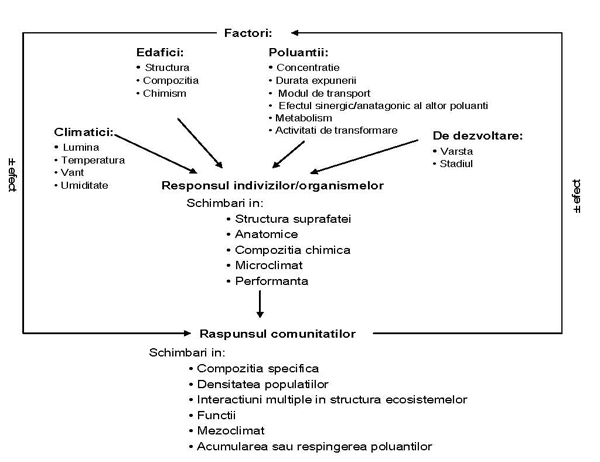
Impactul factorilor de mediu asupra biodiversităţii urbane Marilena Onete Institutul de Biologie Bucureşti, email: m_onete@yahoo.com Principalele probleme cu care se confruntă oamenii acestui secol - schimbările climatice, pierderea biodiversităţii şi creşterea populaţiei umane în special din mediul urban - sunt strâns interconectate (Sukopp et all., 1995). Diversitatea biologică (biodiversitatea) reprezintă variabilitatea organismelor vii incluzând diversitatea specifică, a ecosistemelor (terestre, marine şi alte ecosisteme acvatice), a complexelor ecologice din care ele sunt parte (CBD, 1993). Biodiversitatea este fundaţia pe care este contruită civilizaţia umană. Resursele biologice includ resursele genetice, organisme, populaţii sau orice alte componente ale ecosistemelor care pot fi folosite sau au valoare pentru umanitate (COP, 2002). Aşezările umane, dezvoltate ca mărime şi complexitate de-a lungul timpului, reprezintă nu numai forme importante ale coexistenţei umane dar şi ale relaţiilor dintre om şi mediu. Un oraş este alcătuit dintr-un mozaic de bioTopuri (are structură heterogenă), sediul unor biocenoze în care speciile au cerinţe biologice foarte variate, astfel diversitatea specifică şi densitatea populaţiilor din ariile urbane este mai mare decât a ecosistemelor originare (Sârbu, 1999). Sistemele urbane, complexele industriale sunt sisteme parazite din punct de vedere energetic, depinzand strict de energia şi materia primă din sistemele naturale furnizoare de materie şi energie (Vădineanu, 1998). Imput-ul de materie şi energie pentru menţinerea funcţionalităţii oraşelor este foarte mare. Emisiile solide, lichide şi gazoase sunt cele mai mari output-uri ale oraşelor, care reprezintă imputu-uri pentru zonele adiacente cât şi pentru oraşul însuşi (feed-back pozitiv) (Sukkop et all., 1990) . Factorii care afectează biodiversitatea (inplicit pe cea urbană) sunt multipli, se influentează reciproc, iar răspunsurile diverselor nivele de organizare a vieţii sunt complexe şi interconentate atât în interiorul nivelului, între nivele, cât şi cu factorii abiotici (Figura 1). Oamenii influenţeazǎ mediul lor de viaţǎ introducând şi eliminând specii. Factorii de stres se intensificǎ în oraş (temperaturi crescute, precipitaţii scazute, poluare, etc) şi determinǎ scǎderea rezistenţei organismelor şi eventual moartea lor. Oraşul este considerat ca „insulă de caldură”. Efectul factorilor climatici urbani asupra zonelor verzi este foarte mare.
Fig. 1. Factorii de mediu şi impactul lor asupra biodiversităţii urbane (adaptare după Ashmore, 1998)
Zonele verzi din oraşe, cu cât sunt ecosisteme mai mici şi mai simplificate cu atât sunt mai afectate de factorii de mediu biotici şi abiotici. Cu cât sunt mai complexe şi mai extinse, cu coridoare de tranziţie între ele, influenţează climatul la scară regională. Rata de pierdere a biodiversităţii este fară precedent, ameninţând chiar existenţa vieţii. Menţinerea biodiversităţii este condiţie necesară pentru dezvoltarea durabilă şi constituie una dintre cele mai mari provocări ale lumii moderne (COP, 2002). Bibliografie selectivă: *** COP 6 Decision VI/26, 2002, http://www.cbd.int/decisions/?m=COP-06&id=7200 *** Convention on biological diversity, 1993, http://www.cbd.int/doc/legal/cbd-un-en.pdf Müller N, Knight D., Werner P (Ed.), 2008, Urban biodiversity and design. Implementing the Convention on Biological Diversity in towns and cities, Book of abstracts, Third Conference of the Competence Network Urban Ecology, Erfurt, Germany, 21-24 Mai, http://www.bfn.de/fileadmin/MDB/documents/service/skript229-1.pdf Sukopp H., Hejny S. (Ed.), Kowarik I (Co-Ed.), 1990, Urban ecology. Plants and plant communities in urban environment, SPB Academic Publishing bv., 282 pag. Sukopp H., Numata M., Huber A., 1995, Urban ecology as the basis of urban planning, SPB Academic Publishing bv., The Hague, the Netherland, 218 pag. Vădineanu A., 1998, Dezvoltarea durabilă: Teorie şi practică, Vol. I, Editura Universităţii din Bucureşti, 247 pag. Sârbu C., 1999, Reabilitare urbană şi dezvoltare: o dimensiune principală a tranziţiei socio-economice. Un exemplu de abordare – textura urbană, în: Vădineanu A., Negrei C., Lisievici P. (Ed.), , Dezvoltarea durabilă: Mecanisme şi instrumente, Vol. II, Editura Universităţii din Bucureşti, 298-327.
Carmen Mădălina Cişmaşiu Institutul de Biologie Bucureşti, email: madalinabio@yahoo.com ; carmen.cismasiu@ibiol.ro Cercetarea ştiinţifică s-a axat pe mediile extreme datorită potenţialului pe care îl au microorganismele extremofile în biotehnologie. Studiul ecologiei microbiene a mediilor extreme, bogate în metale, oferă baza pe care s-au dezvoltat tehnologiile de procesare a minereurilor. Cercetarea în domeniul ecologiei mediilor extrem de acide, cu concentraţii crescute de ioni metalici, a relevat că diversitatea microorganismelor acidofile este considerabilă. Aceasta cuprinde în principal ca reprezentanţi Bacteria şi Archaea, precum şi alte microorganisme cu efect sinergic sau antagonic asupra procesului de oxidare a minereurilor sulfurice (Zarnea, 1994 ; Johnson, 1999 ; Lazăr, 2001). Acţiunea bacteriilor acidofile la nivel de ecosistem este foarte complexă: (a) în lanţurile şi reţelele trofice; (b) în limitarea dezvoltării populaţiilor unor anumite organisme; (c) în formarea şi menţinerea structurii solului, esenţial pentru circulaţia apei, aerului şi nutrienţilor; (d) bioindicatori pentru “sănătatea” ecosistemelor (Johnson, 1999; Cişmaşiu şi colab., 2000; Voicu şi colab., 2000; Cişmaşiu, 2008). Dezvoltarea proceselor biotehnologice, bazate pe activitatea microorganismelor acidofile, a demonstrat eficacitatea acestora în îndepărtarea ionilor metalici din efluenţi minieri cu două efecte benefice:
Bacteriile acidofile prezente în drenaje acide de mină şi în solul învecinat acestora pot fi heterotrofe sau chemolitotrofe, fiind implicate în procesele de biosolubilizare, bioacumulare şi biofixare a metalelor. Ele semnalizează apariţia unor modificări negative ale ecosistemelor ce decurg din activitatea unor poluanţi sau alţi factori perturbatori, înainte ca acestea să afecteze organismele mai evoluate). In ultima decadǎ cercetǎrile legate de bioremedierea mediului poluat cu metale grele au luat o mare amploare, bazatǎ pe activitatea microorganismelor acidofile a cǎror diversitate metabolicǎ explicǎ eficienţa îndepǎrtǎrii compuşilor toxici din mediul înconjurǎtor (Rakesh, 1990; Zarnea, 1994; Zarnea şi Dumitru, 1994; Lazǎr, 2001). Accentuarea gradului de poluare a mediului a determinat creşterea interesului cu privire la rezistenţa microbianǎ la ioni metalici şi mai ales asupra extinderii potenţialului de aplicatii biotehnologice acestor microoraganisme. Microorganismele acidofile, cum sunt: bacteriile heterotrofe acidofile şi bacterii chemolitotrofe fier- şi sulf- oxidante, care s-au dovedit rezistente la concentraţii crescute de cupru sunt de interes pentru aplicaţii biotehnologice (biohidrometalurgice) (Voicu şi colab., 2000, Cişmaşiu, 2002). Bacteriile acidofile prezintǎ un interes deosebit în aplicaţii biotehnologice: bioleşiere (biominerit) şi biosorbţie pentru recuperarea unor cantitǎţi apreciabile de metale, în controlul poluǎrii şi îmbunǎţirea combustibililor fosili (Rakesh, 1990, Lazăr, 2001). In urmǎtorul mileniu se preconizeazǎ o extindere pe scarǎ largǎ a bioprocesǎrii minerurilor, cǎt şi a tipurilor de minereu care pot fi astfel preluate. Diferite tulpini, specii sau chiar genuri de microorganisme acidofile pot deveni cei mai importanţi agenţi de oxidare a minereurilor. Dizolvarea reductivǎ a minereurilor conţinǎnd fier feric de cǎtre bacteriile fier-oxidante acidofile poate fi folositǎ la îndepǎrtarea impuritǎţilor de oxizi de fier din depunerile miniere (caolinul, ilmenitul). Alte posibile aplicaţii ale microorganismelor acidofile includ folosirea bacteriilor fier-reducǎtoare pentru bioremedierea controlatǎ a apelor acide reziduale (Johnson, 1999). Biogeotehnologia include biohidrometalurgia (leşierea microbianǎ), bioacumularea metalelor, bioprecipitarea metalelor, biosorbţia metalelor şi desulfurizarea combustibililor fosili. Biohidrometalurgia este un domeniu de mare importanţǎ, cunoscut fiind faptul cǎ peste 20% din producţia mondialǎ de cupru este obţinutǎ din minereuri prin tehnologie microbianǎ. Aceastǎ tehnologie are la bazǎ în principal activitatea microorganismelor chemolitotrofe acidofile din genurile Thiobacillus, Leptospirillum, Sulfolobus, Metallosphaera, care se dezvoltǎ pe baza sulfului şi fierului din mineruri sulfurice, pe care le oxideazǎ în prezenţa oxigenului atmosferic. Alte microorganisme acidofile acumuleazǎ, fixeazǎ si precipitǎ metalele, participînd la formarea de noi zǎcǎminte metalifere (Karavaiko şi colab., 1988; Rakesh, 1990, Lazăr, 2001; Cişmaşiu şi colab., 2007). Activitatea şi selectivitatea proceselor de recuperare a metalelor se datoresc proprietăţilor fiziologice ale microorganismelor, caracteristicilor elementelor, cât şi factorilor fizici şi chimici ai mediului (temperatură, pH), modului de preparare a biomasei, compoziţiei mediului nutritiv, prezenţa diferitelor substanţe organice în apa reziduală, acestea constituind elemente de bază în înţelegerea şi controlarea proceselor microbiologice de îndepărtare a ionilor metalici din ape reziduale industriale (Cişmaşiu, 2003; Toniuc şi colab., 2005; Cişmaşiu, 2006). O importanţă deosebită în utilizarea microorganismelor acidofile în procese de biosorbţie, bioacumulare şi biosolubilizare a metalelor din ape reziduale industriale (în special drenajele miniere acide) o reprezintă rezistenţa acestor microorganisme la concentraţii crescute de ioni metalici existente în mediile respective. Acest fapt oferă posibilitatea utilizării lor eficiente în procesele biotehnologice. Pe de altă parte, studiul rezistenţei microorganismelor acidofile la concentraţii diferite de ioni metalici contribuie la îmbunătăţirea proceselor de biosorbţie, bioacumulare şi biosolubilizare a metalelor şi eliminarea acestora din ape reziduale industriale, prin stabilirea experimentală a influenţei ionilor metalici asupra dezvoltării acestor microorganisme (Cişmaşiu şi colab., 2003; Popea şi colab., 2003; Cişmaşiu, 2005).
POLUAREA APELOR SI METODELE ADECVATE PENTRU DISTRUGEREA POLUANTILOR Petruta Oancea, Tatiana Oncescu Universitatea Bucuresti, Facultatea de Chimie, Catedra de Chimie Fizica
Activitatea socială şi economică are drept efect dispersarea permanentă în mediul înconjurător a unei game variate de substanţe nocive. Decontaminarea apei pune probleme economice şi de logistică de când poluanţii deşi în concentraţii mici sunt prezenţi pe o arie largă. Deversarea apelor uzate menajere sau industriale insuficient epurate, depozitarea necontrolată a unor deşeuri menajere şi industriale precum şi unele activităţi specifice din agricultură conduc la deteriorarea continua a calitaţii apelor de suprafaţă şi chiar a celor subterane. Se stie ca insecticidele sunt compusi cumulativi si toxici care contamineaza mediul acvatic si solul provocand probleme serioase de sanatate oamenilor si celorlalte vietuitoare si plante. În timpurile îndepărtate când omenirea nu era atât de tehnologizată şi deci concentraţia poluanţilor era incomparabil mai mică decât astăzi, decontaminarea era realizată lent de către Natură. Poluanţii erau degradaţi printr-o oxidare blândă cu O2 atmosferic dizolvat în apă, astfel încât se menţinea un nivel acceptabil, nepericulos al acestora. O metodă viabilă de tratament a pesticidelor ar trebui să fie neselectivă, să acţioneze rapid şi să convertească compuşii originali la intermediari mai puţin toxici, biodegradabili sau, ideal, ar trebui să conducă la substanţe anorganice simple ca CO2, H2O, Cl-, NH3 etc.( adică la mineralizare ). Au fost propuse şi testate o serie de strategii pentru a distruge materialele toxice din apă. Posibilele metode includ: îndepărtare fizică, biodegradare, tratament chimic, respectiv fotochimic. În general eliminarea poluanţilor organici din soluţiile apoase necesită una sau mai multe tehnici de tratament primar: oxidarea chimică, desorbţia aerului, extracţii lichid-lichid, adsorbţie, osmoza inversă, ultra filtrare şi tratament biologic. Procesele de oxidare chimică pot fi divizate in două clase: - Tratamente chimice clasice - Procese de Oxidare Avansată (AOP) Această din urmă metodă recurge la fotosensibilizatori, agenţi oxidanţi care sub acţiunea radiaţiilor UV generează radicali cu un potenţial mare de oxidare. Aceşti fotosensibilizatori pot acţiona în sisteme omogene, cum este H2O2 , O3 şi sărurile de fier precum şi în sisteme eterogene cum sunt suspensiile de TiO2. Procedeul H2O2/UV s-a dovedit eficace pentru distrugerea poluanţilor organici ca benzen, toluen, clorbenzen, 2+clorfenol, 2,4-diclorfenol, 2,4,6-triclorfenol, dimetilftalat. Distruge de asemenea şi compuşii alifatici halogenaţi ca 1,2-dibromo, 3-clorpropan, tricloretilena TCE, percloretilena PCE /51-53/. Tehnica AOP a condus la elaborarea unor tehnologii de oxidare avansată AOT folosite astăzi industrial la depoluarea apelor contaminate. Cele mai multe sunt utilizate în America de Nord, în Statele Unite şi Canada . Avantajele tehnologiei UV/H2O2 sunt următoarele: 1. Fotocatalizatorul H2O2 este o substanţă accesibilă, uşor solubilă şi manevrabilă . 2. Expunerea la radiaţiile UV conferă prin radicalii ·OH generaţi o viteză mare de oxidare. 3. Oferă o scară largă de aplicabilitate indiferent de calitatea apei. 4. Dimensiunile echipamentului sunt convenabile. Dacă poluanţii sunt în concentraţii mari sau sunt caracterizaţi printr-o valoare a coeficientului molar de absorbţie mare pentru l < 300 nm, sau dacă au un COD crescut, atunci tratamentul H2O2/UV ca şi cu ozon nu mai este suficient. De aceea s-a făcut apel la compuşii cu fier care de obicei absorb şi în domeniul vizibil în timp ce poluantul nu. Cum în natură scoarţa pământului este bogată în săruri de fier bi şi trivalente iar apele în carboxilat/policarboxilat (citraţi, oxalaţi) se poate conchide că iradierea solară a acestora este responsabilă de: - formarea şi distrugerea H2O2 în mediul natural acvatic - distrugerea şi prevenirea acumulării poluanţilor organici - reciclarea CO2 Dacă sistemul conţinând ioni ferici este iradiat de la început în prezenţă de H2O2 are loc o fotoaccelerare a proceselor. Dar şi la întuneric au loc astfel de reacţii de oxidare în prezenţa sărurilor feroase. De altfel şi sărurile ferice în prezenţa H2O2 la întuneric conduc la formarea de Fe(II) şi apoi au loc aceleaşi secvenţe care stau la baza reactivului Fenton folosit la oxidări în Chimia Organică. Trebuie însă menţionat că aceste procese de oxidare la întuneric nu conduc la mineralizarea completă a poluantului organic. Dacă însă are loc iradierea reactivilor Fenton cu UV/vis, viteza de distrugere a poluantului creşte şi se ajunge până la CO2. Avantajul sistemului ferioxalat-H2O2/UV-vis se datoreşte câtorva factori: - folosirea mai eficientă a lămpii de iradiere deoarece ferioxalatul absoarbe puternic în UV dar şi în domeniul vizibil 250-480 nm; - randamentul cuantic de formare de Fe(II) este mare (1-1,2) în UV şi relativ independent de lungimea de undă; valoarea lui micşorându-se treptat până la 480 nm; având totuşi o valoare acceptabilă. Prin folosirea suspensiilor de semiconductor TiO2 se trece la folosirea fotocatalizatorilor în fază heterogenă. În astfel de cercetări este folosit anatasul ca formă fotochimic activă şi nu rutilul. Sub acţiunea radiaţiilor UV are loc separarea de sarcini cu apariţia electronului în banda de conducţie şi prezenţa golului în banda de valenţă cu o zonă interzisă de 3,2 eV. De aceea sunt eficienţi numai fotonii de energie egală sau mai mare decât aceasta ceea ce corespunde la l £ 385 nm.
REFERINTE
1. J.R. Bolton and Stephen R.Cater., eds., Lewis Publisher, Boca Raton, FL., 1994, pp 467-490. 2. J. R. Bolton, J. E. Valladares, J. P. Zanin, W. J. Cooper, M. G. Nickelson, D. C. Kajdi, T. D. Waiter, C. N. Kurucz. “Figures-of-Merit for Advanced Oxidation Technologies A Comparison of Homogeneous UV/H2O2, Heterogeneous UV/TiO2 and Electron Beam Processes”, J. Adv. Oxid. Technol. 3, 2 (1998) pp. 174-181. 3. Carey, J.H. An introduction to AOP for destruction of organic in wastewater. Water Pollut. Res. J. Can., 1992, 27, 1-21.
IMPACTUL MODIFICARILOR CLIMATICE ASUPRA FUNGILOR Ioana Gomoiu Institutul de Biologie Bucureşti, email: ioana.gomoiu@ibiol.ro
Fungii ocupă un loc important în ecosisteme prin rolul lor de descompunători ai materiei organice de provenienţă vegetală şi animală, în circuitul nutrienţilor dar şi de reglator al numărului de indivizi dintr-o populaţie sensibilă la speciile patogene sau potenţial patogene. La nivel international se constată că domeniului fungilor i se acordă o importanţă mai mică în comparaţie cu celelalte grupuri de organisme, fapt susţinut de următoarele constatări:impactul modificărilor climatice asupra grupelor ecologice şi taxonomice este puţin studiat;organizaţiile guvernamentale şi de conservare acordă atenţie redusă fungilor care sunt organisme diferite de plante şi animale dar trebuiesc protejate iar pentru conservare sunt necesare strategii diferite; în strategiile planificate de protecţie a biodiverităţii fungii sunt foarte rar incluşi; societăţile micologice naţionale şi locale nu există sau sunt puţine la număr şi nu au o politică referitoare la efectul schimbărilor climatice;sunt foarte puţine cercetări în domeniul influenţei schimbărilor climatice asupra fungilor care parazitează plantele de cultură, pădurile, animalele, populaţia umană sau implicaţiile lor în carantină şi bioterorism. Modificările climatice majore includ:schimbări în regimul temperaturii, precipitaţiilor, creşterea frecvenţei şi intensitătii furtunilor (inclusiv a uraganelor) şi a anilor secetoşi , creşterea nivelului de radiaţii UV-B, CO 2 şi a poluării cu azot. Modificările climatice afectează biodiversitatea fungilor şi relaţia patogen-gazdă iar intensitatea acestora este dependentă de acţiunea lor izolată sau simultană;ex: prin creşterea nivelului de CO2 este stimulată fotosinteza deci creşte biomasa vegetală dar prin acţiunea UV-B aceasta se reduce iar sensibilitatea plantelor la fungii patogeni creşte. Efectul modificărilor climatice asupra biodiversitătii fungilor Variaţiile de temperatură şi UV-B acţionează diferit asupra fungilor de pe suprafaţa frunzelor. Astfel speciile care nu posedă pigmenţi melanici în structura pereţilor celulari (hifal şi sporal) sunt mai sensibile decât cele care posedă (Aspergillus niger,Aureobasidium pullulans ). In litiera de Betula pubescens, Quercus robur şi Vaccinium uliginosum care conţine genurile: Cladosporium, Cystodendron, Phoma, Acremonium, Mucor, Truncatella şi Penicillium s-au pus în evidenţă:modificări în structura comunităţilor fungice, reducerea capacităţii de colonizare şi a ratei de descompunere. Deşi incălzirea globală ar trebui să determine creşterea ratei descompunerii acest lucru nu are loc deoarece plantele expuse la UV-B, după senescenţă şi moarte sunt mai greu descompuse ca urmare a efectelor temperaturii şi UV-B la nivelul compoziţiei chimice al peretelui celular precum şi la nivelul materialului genetic (Caldwell et al.,2003). Fungii micorizanţi localizaţi în sol, la nivelul rădăcinilor sunt mai puţin expuşi dar sunt afectaţi de efectul direct al modificărilor climatice asupra plantelor gazdă (din simbioză). Efectul schimbărilor climatice asupra patogenilor plantelor şi bolilor produse Studiul bolilor plantelor presupune analiza componentelor triunghiului:gazdă, patogen, mediu. In general, fungii care produc boli la plante se dezvoltă optim la valori moderate de temperatură. PhyTophtora infestans care atacă tomatele şi cartoful infestează şi se dezvoltă optim în condiţii de umiditate crescută şi la valori de temperatură de 7,2-26,80C. Când valorile ridicate de temperatură se înregistrează mai devreme în zonele temperate, infestarea are loc timpuriu, potenţialul epidemic este crescut iar cantitatea de pesticide folosită este mai mare, ceea ce determină creşterea gradului de poluare la nivelul solului şi freatic. Seceta va determina plantele să reţină apa la nivelul frunzelor ceea ce va favoriza dezvoltarea patogenilor şi răspândirea bolilor. Creşterea nivelului CO2 ar putea determina obţinerea unei cantităţi mai mari de biomasă, aceasta nu va putea fi descompusă pe parcursul iernii dar va reprezenta mediul de multiplicare a patogenilor, astfel încât bolile vor debuta sever primăvara timpuriu .Unele specii patogene sporulează abundent în prezenţa CO2 ceea ce echivalează cu creşterea şanselor de inbolnăvire a plantelor. Puccinia recondita este redusă numeric de valorile scăzute ale temperaturii din timpul iernii dar prin încălzirea globală acest patogen al grâului (în Scoţia) , se extinde. In anul 2050 se estimează creşterea arealului acestui patogen (aproximativ 50% din suprafaţa regatului). Puccinia graminis, patogen al grâului neidentificat în prezent în Marea Britanie şi identificat sporadic în Romania, se va răspândi pe 75% din suprafaţa celor două ţări Fusarium graminearum şi Fusarium avenaceum sunt patogeni ai cerealelor şi producători de mycotoxine. Prin încălzirea globală va creşte riscul apariţiei bolilor, a sintezei micotoxinelor şi a cantităţii de fungicide Efectul modificărilor climatice asupra formării copurilor de fructificaţie la bazidiomicete Cercetările efectuate în herbarele micologice au demonstrat că procesul de fructificare al bazidiomicetelor s-a modificat considerabil în perioada 1940-2006. Astfel, în 1940-1980 întârzierea apariţiei corpurilor de fructifcaţie a fost de maximum 13,3 +/- 1,2 zile iar în perioada 1980-2006, de 12,9 +/-1,2 zile. Intârzirea s-a observat în principal la corpurile de fructificaţie care apar toamna timpuriu şi a constat în 30 zile. Opus, corpurile de fructificaţie care apar toamna târziu au fructificat normal . Astfel, timpul de colectare a ciupercilor comestibile s-a redus considerabil. Prin analiza inregistrărilor temperaturii şi precipitaţiilor s-a constatat:
Implicaţii în starea de sănătate a populaţiei globului Ca urmare a modificărilor climatice va creşte numărul sporilor din aerosporă si implicit a cazurilor de alergie severă (culminând cu astm şi cancer pulmonar). Creşterea umidităţii ca efect al precipitaţiilor abundente favorizează extinderea speciilor Aspergillus flavus şi A.fumigatus cunoscute ca producătoare de micotoxine.Condiţiile adverse(secetă, stres indus, nutrienţi puţini) cresc sensibilitatea plantelor la patogenii producători de micotoxine. HALOBACTERIILE ŞI IMPLICAŢIILE LOR ÎN FORMAREA EVAPORITELOR Mădălin Enache Institutul de Biologie Bucureşti, email: madalin.enache@ibiol.ro
In ultima perioadă, se observă un interes aparte pentru izolarea microorganismelor din evaporite. Din punct de vedere practic, există unele preocupări legate de o potenţială acţiune a comunităţii microbiene halofile viabile din zăcămintele de sare, privind biodeteriorarea unor containere în care sunt depozitate o serie de deşeuri toxice sau nucleare. Astfel de exploatări de sare vechi, în care se găsesc depozitate reziduuri toxice şi nucleare sunt reprezentate de minele din Germania (mina Asse şi mina Bartensleben – în care sunt depozitate materiale radioactive) sau din S.U.A (mina Keller din Texas şi mina Cote Blanche – în care sunt depozitate produse rezultate în urma prelucrării petrolului sau petrol neprelucrat). Pe de altă parte, multe studii au avut ca scop principal înţelegerea originii microorganismelor prezente în depozitele de sare, în particular a celor haloarheene. Intrebarea principală care se naşte se referă la probabilitatea ca aceste halofile să fie descendenţi ai unor populaţii care au rămas captive în cristalul de sare, în timpul perioadelor geologice traversate în urmă cu multe milioane de ani. In cazul în care răspunsul la această întrebare este pozitiv, este necesar să se explice şi să se înţeleagă cum au supravieţuit aceste microorganisme în mediul hipersalin subteran. Descoperirea unor forme de viaţă în evaporite, medii în care pot fi întâlnite condiţii similare cu cele oferite de Marte sau Europa, a crescut interesul pentru cercetările privind identificarea unor forme de viaţă extraterestre. Cu toate că, NaCl este sarea principală în aproape toate apele hipersaline, compoziţia ionică a lacurilor saline continentale poate să difere substanţial în funcţie de geologia zonei care înconjoară lacul. De exemplu, Marea Moartă este bogată în magneziu şi calciu, microbiota fiind astfel influenţată. Salinitatea apei de mare variază uşor pe glob (de exemplu 34 – 36 g/l în Pacific), iar compoziţia ei pare să fi rămas stabilă timp de peste două miliarde de ani, cu toate că asupra acestui subiect există încă o serie de controverse. Majoritatea mediilor hipersaline sunt derivate din evaporarea apei de mare, susţinută de curgerea restrictivă, temperaturi ridicate, un nivel scăzut de precipitaţii, umiditate scăzută şi o viteză ridicată a vântului. O coloană de apă de mare cu o adâncime de circa 1000 m (35 g/l salinitatea) generează după evaporare 14 m evaporit, în majoritate format din halit. Tinând cont de faptul că unele zăcăminete de sare au grosimi mai mari de câteva sute de metri şi că aproape un sfert din masa Pamântului este reprezentată de evaporite, devine evident că mediile hipersaline s-au constituit în adevărate rezervoare pentru evoluţia pe termen lung a microorganismelor adaptate acestor condiţii specifice. Organisme aparţinând celor trei domenii ale lumii vii s-au adaptat şi pot creşte în medii cu conţinut ridicat de sare dar, pe măsură ce concentraţia de sare creşte, diversitatea generală a grupelor fiziologice scade. Unele grupe fiziologice, cum ar fi nitrificatorii, se pare că sunt absente la concentraţii de sare mai mari de 15% (w/v), cel mai probabil pentru că metabolismul dezasimilator are ca rezultat doar o cantitate mică de energie iar costurile extraenergetice necesare pentru producerea soluţilor compatibili conduc la condiţii nefavorabile desfăşurării acestui proces. Intr-un astfel de context nu se poate afirma că mediile hipersaline sunt sărace în nutrienţi. In multe cazuri, evaporarea a avut ca rezultat, pe lângă concentrarea sării şi pe cea a unor nutrienţi. Haloarhea (ordinul Halobacteriales) se caracterizează prin prezenţa unor structuri arheene definitorii cum ar fi lipidele de natură eterică. Ele necesită concentraţii ridicate de sare pentru creştere şi în general, nu se pot dezvolta în medii cu conţinut de sare mai mic de 10% (w/v). Presiunea osmotică a brinei care le înconjoară este contrabalansată prin acumularea ionilor de potasiu în interiorul celulei. Biochimia acestor microorganisme este adaptată la concentraţii ridicate de sare. Diversitatea ecofiziologică a acestor microorganisme este reflectată în cele 27 genuri caracterizate până în prezent. Unele specii necesită valori ridicate de pH pentru creştere iar altele concentraţii mari de magneziu, reflectând astfel adaptarea la mediile în care ele trăiesc. La concentraţii în NaCl între 20% şi saturaţie (circa 32%, w/v) ale mediului, haloarheele devin microorganismele dominante. In astfel de medii, densitatea celulară poate fi atât de ridicată încât brinele se colorează în diverse nuanţe de roşu, favorizând astfel evaporarea apei prin captarea radiaţiei solare. Culoarea roşie este dată de pigmenţii carotenoizi, care protejează celula de efectele luminii ultraviolete. In momentul în care începe precipitarea clorurii de sodiu, multe haloarhee rămân în interiorul unor incluziuni lichide, care pot constitui 2-6% (w/w) din sarea solară proaspăt recoltată. Acest fenomen a fost observat în salternele solare din întreaga lume. Prezenţa haloarheelor afectează cristalizarea in diferite feluri: - creşterea în dimensiune şi a numărului cristalelor - incluziuni lichide mai mari - favorizarea cristalelor dendritice - accelerarea vitezei de cristalizare Haloarheele rămân viabile în incluziunile lichide pentru cel putin şase luni şi se caracterizează prin motilitate timp de mai multe săptămâni. Există raportări în care este descrisă recuperarea de “bacterii roz” viabile din sare solară păstrată timp de patru sau cinci ani. Rolul microorganismelor în diageneza evaporitelor şi supravieţuirea lor pentru timp scurt în interiorul cristalelor a fost evidenţiat în multe publicaţii. Evidenţa pentru supravieţuirea pe termen lung în depozitele de sare este dificil de determinat şi necesită înţelegerea geologiei şi geochimiei evaporitelor precum şi informaţii despre microorganismele izolate. |
||||||||||||||||||||||||
|
© 2008 Institute of Biology. All right reserved. Send mail to sorin.stefanut@ibiol.ro with
questions or comments about this web site. |